1
Master's Thesis / 修士論文
フィブロネクチン濃度勾配を駆動力とする
コラーゲンゲル内EMT細胞の3次元遊走
山口, 陽平
三重大学, 2008.
三重大学大学院工学研究科博士前期課程分子素材工学専攻
http://hdl.handle.net/10076/10886
複写可
修士論文
平成20年度
フィブロネクチン濃度勾配を駆動力とする
コラーゲンゲル内EMT細胞の3次元遁走
∼
L
.∴.-′.
4L
三重大学大学院工学研究科
博士前期課程
分子素材工学専攻
山口
三重大学大学院
陽平
工学研究科
修士論文目次
1.緒言・
・
・
1-1腹膜透析・
・
・
・
・
・
・
・
・
・
・
・
・
●
●
I-2
1-3
・
・
・
・
・
I-1-3
腹膜・
1-1-4
腹膜透析による合併症・
EMT・
・
I
・
・
・
・
・
・
・
・
・
1-2-1
EMTとは・
1-2-2
EMTを誘導する因子・
・
細胞の逓走・
・
・
・
・
・
・
●
●
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
3-1実験系の作成・
・
・
・
・
・P.2
・
・P.3
・
.P.3
・P.5
・
・
・
・
・
・
・
・
・
・
・
・P.7
・
・
・
・
・
-
・
・P.I
・
・
・
・
・
・
I
・
・
・
・
-
・
・
・
I
-
α5β1-integrinと細胞外マトリクスの関係と役割SyndecanとFNの関係と役割-
・
・
・
・
・
-
・
・
・
・
3-I-1ヒト大網由来中皮細胞(ⅡpMC)の培養-
-
・
・
・
・
-
・
・
・
・
・
-
・
-
・P.7
・P.8
・P.10
・
・p.10
・p.ll
-
・
・P.1
・
integrinと細胞外マトリクスの関係と役割-
・
・P.I
・
・
I-4-2.
3.方法・
・
・
FNの構造と機能・
2.目的・
・
I
1-4-I.
1-4-4.
・
I
I-4.フィブロネクチン(FN)とα5β1-integrin・
I-4-3.
・
・
I
・
1-3-1.EMTにおける細胞遊走・
1-3-2.化学走化性因子・
●
・
I
・
・
・
・
・
・
・
●
・
1-ト1腎不全・
1-1-2
連続携行式腹膜透析治療(CAPD)
・
●
-
-
-
・p.12
・p.13
・
・
・
・
・
・p.14
・
・
・
・
・
・P.15
・
・
-
・
・
-
・
-
・
・P.15
-p.15
培地の調整
初代培養の方法
継代培養の方法
3-I-2
ⅡPMC-のTGFIP1誘導・
Growth-
arre
・P.16
st
TGF-β1曝露
3-2
TGF-P曝蕗による細胞の形態変化の動画(及び静止画)撮
影・
・
3-3
・
・
・P.16
TGF-P暴轟によるSnailとE-cadherin、
・
mRNAの経時変化の測定・
・
MMP-2
α5-integrin,
・
・
・
・
・
・
・
・
・
・
3-311
AGPC法によるmRNAの抽出と定量・
3-312
mRNAの逆転写によるcDNAの作成とpCRによるSnai1、EICadherin、
・
・
・
・
.
-
α5-integrin及びMMP12DNAの増幅31313.電気泳動とバンドの定量・
・
・
・
・
・
・
・
I
・
-
・
・
・
I
-
・
・
・p.17
・
・
-
・
・
・P.17
-p.17
・
・
・
・p.1S
EMT細胞の三次元細胞遊走評
3-4.化学走化性因子を用いた
価・
・
・
・
・
・P.19
評価モデルの作成
3-4-1.FNおよびBSAを用いた三次元遊走モデルの作成・
3-4-2.
・
・
・
I
・p.19
p.20
RGDS阻害剤によるFN存在下での三次元遊走モデルの作成-
3-4-3.遊走細胞数の計測・
3-4-4.細胞増殖測定・
3-5.
・
I
・
.
・
・
・
・
・
I
・
・
・
・
・
・
・
・
.
I
・
I
・
・
・
I
・
・
・
・
・
・
・
・
・
・
I
・
・
・p.20
・
.p.20
TGF-P1誘導時のFN添加及びRGDS阻害によるα-SMAの経時
測定・
・
・
・
・
3-5-I.タンパク質の抽出とBCA法による定量・
3-5-2.
SDS-PAGEによるタンパク質の分離・
I
-
3-5-3.膜-の転写(ウェスタンブロツティング)
三重大学大学院
・
工学研究科
・
・
・
・
・
・
・
・
・
・P.2l
・
・P.21
・
・
・P.21
・P.22
3-5-4.抗体反応とバンドの定量・
4.結果・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
.
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
4-1.TGF-β1誘導による細胞の形態変化の観察4-2.TGF-β1誘導による細胞の遊走距離の測定・
4-3.
TGF-β
4-3-1.SnailmRNA.
4-3-2.
・
E-cadherin
4-3-3.
a
4-3-4.
A4MP-2
・
1誘導によるSnai1及びE-cadherin、
MMP-2mRNAの経時的測定・
・
・
・
・
・
・
・
・
・P.22
・P.23
・
・
・
・
・p.23
・
・
・
・
・p.24
α5-integrin
・
・
・
・
・
・
,
・p.25
・
・
・
mRNA
5-integrln
・
・
・
P.25
・
P.26
I
P.27
●
・
mRNA
・
mRNA
・
P2S
・
4-4.化学走化性因子を用いたEMT細胞の三次元細胞逓走評
価・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
4-411.FNを用いた遊走細胞数の計測・
I
・
4-4-2.BSAを用いた遊走細胞数の計測・
4-4-3.
FNを用い,
TGF-β
・
・
-
I
・
・
・
・
・
I
・
・
-
・
・
I
・
I
・
・
・
・
・
・p.29
・
-
-p.29
・
・
・
・
・
・
・
I
・
・
・
・
・
・
・
・
I
.
・P.29
・p.30
・
・
・
・
・p.31
1誘導時のFN添加及びRGI)S阻害によるα-SMAの
轟時的測定・
5.考察・
・
RGDSにより阻害した際の遊走細胞数の計測-
4-4-4.細胞増殖試験・
4-5.
・
・
・
・
・
・
・
・
・
・・
・
・
・
・
・
・
・
・
5-I.腹膜透析によるTGF-β1産生機序・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・p.32
・p.33
・
・p.33
β1によるSnail
5-2. TGF-
減少メカニズムとその役割・
5-3.
・
・
・
・・
・
・・
・
5-4.FN阻害剤としてのRGI)Sの妥当性・
6.##・
・
・
・
・
・
9.謝辞・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
PENDIX・
・
・
・
・
・
・
mRNAおよ
・
・
・
・
・
・
・
・
・
・
・
・
-
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
I
A13.AGPC法プロトコル.
A-4.RNA定量プロトコル・
・
・
I
・
・
・
・
I
・
・
・
I
三重大学大学院
・
・
・
・
・
・
・
・
・
・
・
.
・
・
・
・
t学研究科
・
・
・
・
・
・
・
・
・
・
・
・
・
・p.49
・
・
・
・
・
・
・
・
・
・
・
・p.43
・p.44
・
・
・
・
・
・
・
I
・P.41
・
・
・
・P.40
・
・
・
・p.39
・
・
・
・p.36
・
・
・
A-2.細胞の2次元遊走距離の測定(ImageJの使い方)
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
・
A-1.細胞培養プロトコル・
A-5.RT-PCRプロトコル・
・・p.34
・
・
・
・
・
・
7.今後の展望・
8.参考文献・
・
MMP-2
びα-SMAの遊走における役割・・
10.AP
・
EMT誘導HPMCにおけるα5-imtegrin,
5-5.総括・
mRNAの
mRNA発現に伴うE-cadherin
・
・
・
・
・
・P.50
・
・
・P.50
・
・P.56
・
・P.60
・
・P.63
I
・P.65
A-6.ウェスタンプロット法・
-
・
A-7.三次元細胞遊走モデルの作製
A-S.細胞増殖試験・
・
B-I.滅菌について・
B-2.PrimerList・
I
・
・
・
・
.
I
・
・
I
-
・
I
・
・・
・
・
'
・
・
・
・
'
・
・
・
B-3.RT/PCRprogram'
B-4.統計処理・
I
・P.96
・
・
・
'
・
・P.82
I
I
-
・
P.102
P.105
P.107
I
P.108
I
P.Ilo
1.緒言
1-1.腎不全と腹膜透析
I-111.腎不全【l}]
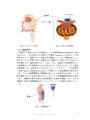
腎臓は尿管や血管,実質(皮質及び髄質)などから構成されている
(Fig.1-1、 1-2)。実質は主に糸球体とボウマン嚢で構成される腎小体とそ
れに続く尿細管で構成され、その構成単位はネフロンと呼ばれる(Fig.113,
1-4)。このネフロンといわれる組織が、腎臓の機能である体内の不要な老
廃物や水分の排除,電解質(ナトリウム,マグネシウム、リンなど)やpH
の調節などを行っている。また尿の排惟によって、タンパク質代謝-産物(尿
素、クレアチニンなど)の除去や血液の浸透圧・pH調整、細胞外液量の調節
が行われる。しかし免疫系の異常や薬に対するアレルギー、高血圧、糖尿
病などが原因で腎臓が十分に機能しなくなるo
このような状態になること
を腎不全といい,週単位で急激に起こる可逆性の急性腎不全と、年単位で
進行する不可逆件の慢性腎不全に大別される。また腎不全は腎臓の機能が
50%を下回ることをいい、
10%をきると末期腎不全とよび,何らかの対処
療法を受けなければ死に至るo
左腎臓
勝朕
ー一---ー
Fig.1-1泌尿器系器官の位置
Fig.1-2左腎の断面
メサンギクム
尿細管
ボウマン嚢
Fig.1-3ネフロン単位
Fig.1-4腎小体の構造
Ill-2.腹膜透析P)
末期腎不全患者に対する対症療法の一つに腹膜透析(peritonealDialysis :略称
pD)があるo
これは患者自身の腹腔内に高濃度D-glucoseを主成分とした透
析液を注入し,腹膜を介して拡散と限外漉過によって水・老廃物(尿として
排出されるべき物質)の除去,電解質・pHの調節を行う療法である(Fig.ト5)。
血液透析に比べ通院回数が月に1-2回と少なく、連続的な老廃物除去によ
る循環器系-の負担軽減から社会生活を営む上で有利である。しかし腹膜
透析療法では、透析液中に含まれるグルコースやグルコース分解産物
(GDP :glりCOSedegradation products)の影響により,溶質透過性克進や除水
不全、さらには腹膜線維症や腹膜硬化症といった合併症を引き起こし、治
療を継続できない症例も報告されている【4]oまた長期腹膜透析患者の増加
に伴い、腹膜の形態学的変化や溶質透過性の克進等が問題となり、その早
期診断と同時に腹膜機能劣化のメカニズムについての解明が急務となって
いるo
)T
1
Fig.
∫
115腹膜透析Pl
弔ノニノil:)(;:: r;;E
1
JI:LJf'')iI:
1-1-3.腹膜組織の分布と構造【5]
腹膜は腹壁の内面を覆う壁側腹膜と腹部臓器(肝臓、肺臓、牌臓、腎臓)
とその間膜(大網,胃、腸間膜)を覆う臓壁腹膜からなる衆膜組織で、表面
は多くの微繊毛を持つ一層の中皮細胞で覆われており、基底膜下(問質)に
は弾性繊維、勝原繊維、毛細血管、肥満細胞などで支持されている複合的
な膜と定義されている(Fig.1-6).腹膜の面積は、成人では1.7
m2-2.2
m2で
ある。腹膜の厚さは50-150pmで、壁側腹膜と臓側腹膜に囲まれた空間を
腹腔といい、ここに腹膜透析液が貯留される。
(microvifli, MV
;
junctional complex,
body.
celt,
cilium,
JC
LB..
M
;
elastic
fibroblast,
E;
;endothe暮ium,
basement
membrane.
C
;
;
lame‖ar
lamina.
EL
; mast
F ; Pericyte,
capi一一ary, CAP
P
;
BM)
Fig.1-6正常腹膜の形態学的模式図
transition
(EMT)
Ill-4.腹膜透析とEpithelial to mesenchymal
腹膜透析の長期化により,腹膜炎や腹腔内出血が繰り返し引き起こされ
る。さらには腹膜硬化症や腹膜線維症を引き起こすこともある。腹膜線維
症は問質内に存在する細胞が,コラーゲンなどの細胞外マトリックスを過
産生する事により問質が線維化することで、さらなる腹膜機能の低下をも
たらす【6).今まではこのような腹膜組織の機能・構造変化は、主に腹膜問質
線維芽細胞によって生じているものと考えられてきたが、上皮細胞マーカ
ータンパク質サイトケラチン陽性細胞が問質内で観察されたことから、上
皮細胞の関与も議論され始めている【7】 (Fig.1-7茶色:サイトケラチン陽性
細胞)。最近では線維症に関係する細胞の起源として以下の三つの説が提案
されている。
3
..T1
、_
:
.一ヽ
I..
_/
「
1
・・■
ヽ
1
、1
I
-i:i
、L(,-1・T:.・'!・
、\l
t:
'i:.vfi.;.
●J
I
ヽ
.thlk
I
tふヽ
1
:、
、ヽ・
_I_I
:it'.i態
?.*.丁
、1・・,
1
.
l
,、ト:it
I
▲
Fig,1-7問質内での上皮細胞の観察
仮説(1)
Ⅰ℡MCが問質-遊走した細胞由来【7・S]
透析液の高浸透圧や酸性、加熱滅菌時に生じるGDPs
degradation
HpMCはEMT
(glucose
products)の影響で、 HPMCは剥離し遊走する。このとき
(上皮間菓系転換)を引き起こして、細胞間接着を失う
ことで上皮細胞の極性を失い、問質層-と遁走する。
仮説(2)循環白血球由来r9J
血液中を循環している白血球が、組織障害部位へ遊走することによ
る影響。
仮説(3)もともと問質に存在する線維芽細胞由来【10】
組織傷害が生じた際に、傷害部位-と遊走し増殖する事で療痕形成
を促進し、組織修復を行う線維芽細胞によって引き起こされる。この
細胞は器官形成後に未分化のまま残った間英系細胞であると考えられ
ているo
:.電人乍人rLP:院
T_/?:研究科
1-2. EMT
l-ユー1.EMTとは
1-2-1-1.
EMTの機能
EMTは上皮系細胞が間英系の細胞に形質変化することで、細胞間接着
の減少やそれに伴い細胞極性を損失する。すなわち上皮細胞の極性を失
うことで、線維芽様細胞-と形質変化する。形質変化した上皮細胞は、
細胞遊走能・浸潤能が大きく増加し、また細胞外マトリックス産生も活
性化される【6].上皮細胞のこのような現象は、傷害修復過程や初期腔発
生における原腸陥入,神経提細胞の運動や器官形成過程,特に心臓や腎
臓、口蓋形成での重要性がこれまでに明らかとなっている【11,12】。また癌
細胞の浸潤や線維症との関連も示唆されている【10】。
I-2-1-2.EMTの指標
上皮系細胞が間菓系細胞に形質変化するとき、構造的・機能的に変化
するため様々な分子を再構成する必要がある。上皮細胞で持っていた
mRNAやタンパク質は維持または減少し、間菓系細胞がもつ特有の
EMTを生じたときに変化する分子
mRNAやタンパク質を発現していく。
マーカーを以下の表にまとめた(table.1-1,1-2)o
Table.1-1
EMTによりup-regulateされる分子
up-regulate
される分子
Snail
α-SMA
機能
転写因子の一つで細胞間接着分子を減少させる○
細胞骨格を形成するタンパク質o
Fibronectin(FN) 細胞接着や遊走に関与する細胞外マトリックスo
Co11喝enⅠ/Ⅲ
ビメンチン
MMP-2/9(57]
問質の主な構成成分である細胞外マトリックスo
開業系細胞の中間系フィラメントの構成成分o
主にrV型コラーゲンを分解する酵素o
5
Table.1-2
EMTによりdown-regulateされる分子
down-regulate
される分子
E-cadberin
Occludins
ZO-1
Desmoplakin
サイトケラチン
機能
Adherencejunctionを構成するタンパク質o
tightjunctionを構成するタンパク質o
occludinsの裏打ちタンパク質o
デスモソ-ムの裏打ちタンパク質o
上皮細胞の中間系フィラメントの構成成分o
E-cadberinやoccludins、
ZO-1、
desmoplakinは細胞間接着に関与するタ
ンパク質であり、 EMTを起こしたときに細胞間接着が解離するために減
少する【13】。
snailは遺伝子レベルで細胞間接着をコントロールする転写因
子である【8]. α-sMAは細胞骨格を形成するタンパク質で、形態変化に伴
い発現する【14】。
FNやcoll喝enは間菓系細胞が産生する主な細胞外マトリ
MMP-2/9
(matrix
ックスで、形質変化することで産生する'ようになる。
metalloproteinase)【57】は細胞外マトリックス分解酵素で主に基底膜の破壊
を破壊し、問質-と遊走し始める【15】。サイトケラチンは上皮細胞、ビメ
ンチンは間菓系細胞に特有の中間径フィラメントの構成成分であり、上
皮細胞の特性を失い、開業系の特性を得るために、それぞれ変化する【101。
これらの分子マーカーを測定する事により、
EMTが生じたかどうかの判
断が可能となる。
1-ユー2.EMTを誘導する因子
腹膜線維症と関連したEMTは、多くのgrowthfactorやサイトカイン、ホ
ルモンなどの細胞外物質によって制御されている。現在報告されている
EMTを誘導する因子を示した(table.1-3)oその中でもTGFgrowth
β 1 (transforming
factor)は様々な病的状態においてEMTを誘導し、問質線維症を導
く因子として特定されており、
EMTを誘導する因子としては最も研究が盛
んに行われている。腹膜組織においても炎症時には過剰に産生することが
報告されている【16・17]。またTGF-β lだけでなく複数の因子を組み合わせる
事により、相乗効果でよりEMTが進行することも報告されており、組織中
では様々な因子が複雑に相互作用している【15】。
I-.重大学大学院
[二号研究科
6
Table. 1-3
EMT誘導因子
分子
TGF-β1
(transforminggr.owthfactDr)
特徴.機能
Reference
多くの細胞に対して強い増殖抑
8,15
制作用を示すo発生や分化にも
18,19
関与するo
EGF
(epidermalgrowthfactx)r)
細胞増殖や遊走、細胞外盲トリ
ツクス産生など様々な機能を持
15,18
つo
FGF-2
(fibroblastgrowthfactor)
線維芽細胞をはじめとして多く
の細胞に対して増殖、分化を促
20
進させるo
ⅠL-1
サイトカインやケモカイン産生
(interleukin)
を増加させる炎症性サイトカイ
21
ンo
AGEs
糖からメイラード反応を経て形
(adVancedglycationend
成されるo老化や糖尿病などに
products)
関与し、体内では非常に分解さ
22
叫こくい.
TypeⅠcム11agen
PDGF
問質の主要な構成成分o
18
腔発生や創傷治癒、炎症時など
(plateletde血ed
様々な状況に関与する.血′ト板
growthfactx)r)
や単球などにより産生されるo
23
1-3.細胞蕃走
1-3_I.
EMTにおける細胞遊走(10}4)
細胞遊走は様々な状況で起こる細胞運動である。例えばEMT時の細胞遊
走や癌細胞の遊走(転移)、白血球の炎症部位-の遊走、傷害修復時におけ
る細胞遊走などがある。ここでは特にEMTにおける細胞遊走を示す。
EMT
が生じてから細胞が遊走するまでに、大きく分けて3段階になる。その模
式図を下に示す(Fig.1-8)0
STEPl
まず初めに細胞間接着を構成するtight
adberence
junction (occludinsやzo-1)や
junction (E-cadherin)の減少によって、細胞間接着が失われる。
さらに上皮細胞のapicalに存在する微繊毛を失うことにより、細胞極性を
7
失うo
STEP2
細胞間接着を失った事により、形態保持が困難になり細胞骨格を形成し
ているα-SMAを再構成する事で形態変化し始めるo
またこのときα-SMA
というストレスファイバーの形成だけでなく,仮足や接着斑を形成するこ
とで、細胞は前後の極性を獲得して遊走し始めるo
STEP3
最終段階として細胞外マトリックス分解酵素MMPを産生する事により
基底膜を破壊し、さらには問質を構成しているI型collagenを分解しなが
ら浸潤していくD
E]句歴=〕嘩ゝ
ーこ卓こゝ
間蕉
STEP
1
STEP
・細胞間接着の解離
apical・basal極性の欠如
=コ
・
Fig,I-8
基底憐
2
STEP
・細胞骨格の再構成
・立走
3
・基底膜破壊
・痩潤
EMTによる細胞遊走
113-2.化学走化性因子
化学走化性因子はEMT細胞を遊走させるだけでなく、癌細胞の遊走や傷
害修復時における上皮細胞の遊走、炎症時における炎症細胞・線維芽細胞
の遊走など様々な状況において機能する。細胞は様々な化学走化性因子が
存在する中で,レセプターを介して遊走のシグナルを細胞内に伝え,より
強い機能を持った因子を認識し,方向性を持って目的とする部位-と遊走
すると考えられている【25lo化学走化性因子として考えられている因子及び
それに対するレセプターを以下にまとめた(table.1-4)o
:.
F!:ノこ,i;i-人号院
r_予研究・fltト
Table.1-4化学走化性因子
分子
FN
機能
レセプター
Integrina5B1
Reference
主に線維芽細胞が産生する、接
(丘bronectin) ⅠntegrinαⅤβ1 着や遊走に関与する細胞外マト
19,26
27,28
リックスo
PDGF
PDGFR1
腔発生や創傷治癒、炎症時など
PDGFR2
様々な状況に関与する.血′ト板
29
や単球などにより産生されるo
IL-8
GPCR
好中球の遊走に関与するケモカ
30
インo
31
多くの細胞に対して強い増殖抑
TGF-β
TGFR
制作用を示すo発生や分化にも
15,18
関与するo
線維芽細胞をはじめとして多く
FGF
EGF
FGFR1-4
HERl/ErB1
の細胞に対して増殖、分化を促
20
進させ争.
細胞増殖や遊走、細胞外マトリ
15
ツクス産生など様々な機能を持
つb
VEGFR1-3
透過性克進因子として機能す
33
るo
endothelia1
growth
32
主に血管内皮細胞の増殖や血脊
VEGF
(Vascular
18
factor)
HA
(hyaluroma皿)
CD44
REAMM
グリコサミノグリカンの一種で
34
様々な細胞動態に関与するo
上記以外にも化学走化性因子としてはケモカインファミリーがある。ケ
モカインとは細胞遊走を主要な作用とするサイトカインの一群であり、炎
症反応、免疫応答などの生命維持活動に極めて重要な役割を担っている。
ヒトでは約45種類のケモカインが同定されている。
9
I-itLtegrin
1-4.フィプロネクチン(F叩とa5β
141.
FNの構造とその機能
FNの役割は細胞接着性の糖タンパクで、腫形成、創傷治癒、ガン転移や
線維化、血栓形成など多くの役割を果たしている。
FNは通常では肝臓で合
成され、可溶性のタンパク質として血簾中に高濃度で存在しており、また
創傷時及び止血時においては局所的に多量に存在するo
FNの構造は、分子量約40-50kDaの糖タンパク質であり,
プOTnI、
Ⅲ、
3種類のタイ
Ⅲ)の繰り返しモジュールで構成されており、
2本のよく似た
サブユニットがジスルフィド結合でつながった構造をしている
r54]f55]G;ig.
1 9)D
152][53]
-
RGDルーープ
12--,壬J789
1
-白i6∵丁-T;1害
ヘパIJン脚位Ⅱ
a5B
l11ntegrhl脚位
COOH
】
l
&Ⅱ
ヘパリン持合軸Ⅰ
Le&a rld
L・・.FnI
FrIZ It
FnI
1
II 2
I 3
Fnェt
FnI
II
FBI
115
4
-
LCPGSSIPPYWET
PLSPtI仙1朋肝ZrrC
a
Ftl=tZ
FELェZI?
・
mOL7DL5FYDエTZ)a
-
-
耳工E)ⅩP神崎V7BVODR甘IDRPXWVt)VD-
EhI
II12
ÅIPAZITD叩T-
FJ)I
TZ
13
‡ⅣS FP捌VTDA7ET-
14
AZZ)APSNFLA一丁P7T-
T -PJIJJ13
-
-YGI吋ZTV7mm
・】)8EQSL一も吉TSC℡r
SI▼WBRPQ・APITGY)IVr8PSVE-Q5S7ELN上℡AF卯ⅦP叩m7rrAYEE・
9AmY7mIFVRP-C耶UZ5脚TQLgI棚AV8エー
mVm柑-AQユ7GmTVGLTRR
-CCPlq耶BvBEyPu叫壬ASmELVAIEG
TZVZTmP・
,
-
・
-
-
91和一p8
・
-
-PェQR71tPDVtSYTrT6LQ仰Yt工Yt,YtLZD
-
PfZmRpRPmTZ一札tPtiTmtYV7AJJtF・
ZVW
EE?LCP珊57aL7ZJTA7R匂A77n
-7ALI邪-ムー呈J工ゆZ且TyCLRDVP
I
CE)ESLP'n't,W
QE】望汀PLRGIQKT
-1EESPuI閉脚
88ヱSCPLVPAV7
1DKESCPZ'rO℡OgT
TLTSIPAQQVYTTt.I
・
St'LVSqPP・RAR)TGYIIⅧPBSP
Qm紺=FQDT8Erll蜘Vm-
I
I
・SSPmrRVW-GP7KTX7^CPJPEmZ叫PW8VYAqWI
SZmアE甜-○¢qVSRYm8gPEDO:皿FPAf瓶TAJ:脚0肌V8VV▲L皿5L$1PPZI一耶t'TmⅦPE正℡
OPm工WZ)8B8VnBmTZLrLrVmAZJD-
-
一杓ES耶T℡
(;qERDAPTVW
.Dt一ざVPTDST一)P
SZ)さmPOTmSVSm
-Tm抽pさrDL7tqLYRYSPVn7Z:SF "
rAP
IRITITQTRT棚田アS-Q7LPRm甜SZTt,T札TPtMSl▼AZJm
I
pqI脚5TGCtALS
-
-
-7Zn紙7t・TFTIT6F叩▲VP▲耶・
且U朋SOlmT-
-1q7aS7mQQET
O&EBXPLT棚T
I
GW一mSZ)BBSrVVS6L℡ymm℡19VJJUI
-AFRIGP柵pSく狩=
VLTV耶R柑pD】T8Tm777P¶脚5L)叩SSC7WPQLrYtNSVY7VmE)エQtJLNW8T工Ⅰ8YnTm▲CE18IPIPDF'VDS9Vm朋f◎IpYt)=8V17t一工甘・
EZWABJJSI
I IED▲
Ft) ⅠⅠI 15
A
AVPPmul円甘IOPD・
FnI
FtIェT=
・
I
CLDSP℡OIt)F昏DZTAJ7・
I l1
FnェZ
・・・ⅡCS
-
入相APpDPTVtPDTTVP5PRDLQm-和肘TmKLDAPTNLq2VmB-
Fn7
FtLIIZ≡Dn
●
・・・FhⅡ
SQPVm=TZFTPSQPFg江PIqN7nZI9pE;甘Ⅰ3RTZIR甘RPRFS
PCNByTIX耶叩TちGQL=SIqQI
-VQ3tmT7
SPL'VATSTSmITASSFVVgyVSAB・DTV8¢ア和E'S耶-DEpQYLDLPS℡ATSV7TI皿mTCエ9E-
FTII工エ6
T17
●.・・hⅡ●
・
-
-且一札SSFVVIDAST
・耶ZSEPL工6紐T
Oq皿ⅩⅤ
-
ODRTTIDLTEZ)EqYSIG和PDTETEVSLISよFEGD-XSSNAXm
RGDS
1
榊:I:
;
E a
Fig.I-9
A
G
F C
D
FNの構造
三重人'1f:人乍院
10
_1二乍研7モ糾
前記の図は、
FNの模式図および各モジュールのシークエンス(FnⅢリガ
ンドのみを示す)とそのモジュールの模式図を示したものになる【55]。
各モジュールは模式図に示してあるように折りたたまれている形をして
おり(左下図)、 N末端から始まりC末端-とつながっている(A⇒B⇒
-
I
Gというフラグメント)。この模式図の緑色で示された部位は,モジュール
の後ろ,赤色で示された部位は手前にあることを示しており,各シークエ
またFNの構造はシークエンス
ンスにそれぞれ色分けで表示をしているo
内の緑(A、
B、
D、
E),赤(c、
G、
F)でそれぞれβシートを形成しており、
色づけされていないシークエンスでループ構造となっている。
このような構造をしているFNにおいてもっとも主要な役割を果たして
いる部位が、リガンドFnⅢの部位である。とくにFnⅢ4-5及びFnⅢ12-14
は-パリン結合部位として細胞表面のプロテオグリカンO'G)と作用し、ま
たFnⅢ7-10特にFnⅢ10に含まれているFNにおいて最も特徴的なループ棉
造をしているRGDSr59](Arginine-Glycine-Aspartic acid-Serine)はintegrinと作
用し、細胞遊走に関与するシグナル伝達を行うことが知られている。
1-4-2.
iAtegrinと細胞外マトリクスの関係と役割
細胞はintegrinを介し様々な細胞外マトリクスと結合する。インテグリン
は140-200kDaのα鎖と90-110kDaのβ鎖が1:1の-テロニ量体で、現在ま
でにα鎖18種類、
β鎖8種類の24種類が確認されている。作用する細胞
外マトリクスはα鎖及びβ鎖の組み合わせによって変化し、
RGDS受容体、
コラーゲン受容体、ラミニン受容体、白血球特異的受容体の5種類によっ
て分類されるO7ig,1-10)o
このような特徴を持ったinteBrinは対応する細胞外マトリクスによって
細胞の増殖、接着および遊走に関与することが知られている。
(
,
\
β4
・
・
-
・collagen
RGD
receptor
receptol
!
-
・Laminin
rECePtOr
-白血球特異receptor
・X・α9β 1はテネイシン・Cと持合する
Fig.1110
integrinとECMの相関図
ll
このようにintegrinは様々なECMと接着することができる。
I-4-3.
α5β 1-integrjnとFNの関係と役割
FNと結合するintegrinはRGDを認識,結合できるサブユニットを持っ
ているQなかでも、もっとも一般的なものがα5β
1-integrinであるo
integrin
は、
α鎖、 β鎖の-テロニ量体で、それぞれのサブユニットの違いにより
名前をつけられているものの、基本的な構造は同じである
RGDS
-I-.i-==-
-.---
----・=pIRGD・-
e.,...d.,,...d亨司
α鵜一日Iahlltrp+d4hdbll
EF仙4typ●d○ft山一lniSYLふY8YAW>eF軸tX}TEEFYA4YPMnNLT
…t●d4rIIJdr)
β1執lllB○LlllE4Fd●bldTLl
S-a
■■LiE事I:EWPIDL.YYLME)はY8MKDDLDIV仰
Fig.1-ll
この図はα5β
integrinとFNの接着
1-integrinの模式図を示したものである.この模式図につい
て説明するとintegrinは膜貫通型の受容体(左上図)で、
を持っているo
α鎖にCa2'結合部位
この部位がCa2'と結合することでα鎖の一部が正の電荷を
帯び、これによりFNのFnⅢ10に存在するRGDと結合し(右上図)、細胞膜
下のチロシンをリン酸化、シグナル伝達が起こると考えられている。この
RGDと結合する部位はα鎖の方はEF-hand
type
domainといわれ、
β垂削ま
DDL配列と呼ばれている(下図)o
このようにα5β1-integrinはFNのRGDS領域と結合し、チロシンをリン
酸化しシグナル伝達を行っている。このようにFNはintegrinと作用するこ
tT・.;大Jj:1こ■f:【ゝ′:-ir √、;モ桝7E不ご1
12
とがしられているが、最近の研究からsyndecanといわれるレセプターとも
結合することが明らかになってきているo
1-414.
SyndecanとFNの関係と役割
Syndecanは細胞表面に存在するプロテオグリカンで、
テオグリカン¢Ieparansulphate
-パラン硫酸プロ
proteoglycan:HSPG)とコンドロイチン硫酸プ
ロテオグリカン(Chondroitin surface proteog】ycan.・CSPG)の対で構成されてい
る【53][54](Fig.1-ll)。
∝謁t宅ミ∃
り
チE)シ
.+
し
(
0
FbZ
I I4
PhI
I =5
TVPSPRZ)L4mVm岬PVmB
Sulphate
Chondroitin乱rrace
chain
mrmPP耳-S^VTCyRVZ)V19VIZLP
I
-
Heparan
7VLⅥmpA-AOZTGl'且LT/SLTRR-
-
I
chaln
GBEGCRLPTSRETrFAEV76LSPmFXVPAVS丘・
00P叫Y7PTQP8VSKYPLqPASE円サEZmIXO-
Fュz Z王12
▲IPAJr7DLmVm-
FELT IT 13
Pzz== ZZ・l
月YSPPF(芦pIRVVD^7TT-
-TI11引柁tEITFP王7W叩^VPAZq;・
-GPJqKJM5SSVW脚SYYALED
-O一打CRTZmVRSYV77Ct々ⅦYt7YE'YtZJq)
A=Z)AP皿Jru77W-
-BLLⅦ押・11ヱr7WJJg7ZXPGSP
-P土帥B抑Pqy7E17=W仰TZ7YM・
I
SLSL叩e-ZWQL76YmV7Y芯ET
Fig, 1112
GRBSXPLT^OQTI
・一浪qESPM¢Vm
-
-
-一TL7SRP即l止
-芯LRSSPVYZt)LET
-
・JFD書SBPZ'1GW
Syndecanの構造(上図)とFNの構造(下図)
このような構造を持っているSyndecanは、
ロイチン硫酸鎖が、
-
-パラン硫酸もしくはコンド
FNのFnⅢ4-5(YRLTVGLTRシークエンス)またはFnⅢ
12-14内のfmS-1(Heparin-binding・site・1:図中イエローマーカー部位)または
f丑IS-2といわれる-パリン結合部位と結合することで、細胞内のチロシン
部位が活性化し、シグナル伝達を引き起こす【64]oこのsymdecanと結合する
FN
の-パリン結合部位について詳しく説明すると、
FnⅢ4-5
の
YRLTVGLTRやFnⅢ12114内に存在するHBS-1(下nⅢ13内の黄色の部位)も
しくはHBS-2(FnⅢ14内の黄色の部位)は、 AZIEやCDGF(Fig.1-9左下図)と
いった同一のβシート上に存在し、
syndecanと結合している。
以上のようにFNはα5βトintegrinもしくはSyndecanを介することで遊
走に関与することが知られており、
EMT誘導により起こる細胞の遊走にも
関与が考えられる。最近の研究でもガンの浸潤の原因にFNが関与してい
るのではないかと考えられている。
13
2.目的
HPMCのEMTは腹膜線維症に関係していると考えられているが、その詳し
いメカニズムはまだ解明されていない。特にEMTを起こしたHPMCがどの
ように問質-と遊走するのかはわかっておらず、そのメカニズムを解明でき
れば、腹膜線維症の予防や治療法の開発に貢献できる。
そこで「炎症等によって問質中に生じた化学走化性因子の濃度勾配に対L
て、
EMTを起こした細胞が遊走する」という仮説を立てた。
この仮説を基に、
(1) TGF-β 1曝露によるHPMCの形態変化の観察及びmRNA測定からの
EMT誘導の確認
(2)EMT誘導HPMCが化学走化性因子の濃度勾配を認識して遊走すること
をin vitro実験系で実証
(3)EMT誘導HpMCの遊走の原因を証明
(4)またその証明のために従来の三次元遊走評価法をより改良した新しい三
次元細胞遊走装置を試作
以上の四つを目的とした。
三重大学大学院
工学研究科
14
3.方法
実験の概要
具体的な研究項目は以下に記す。
Ⅰ. EMT誘導確認実験
①
TGF-
β 1曝露による腹膜中皮細胞(Ⅰ廿MC)の形態変化の動画観察
② TGF-β 1曝露によるHPMCのSnail及びE-cadherin
mRNAの発現測定
Ⅲ.三次元遊走実験
③化学走化性因子FNに対するレセプターα51integrin
mRNA及び
collagen分解酵素MMP-2
mRNAの発現の測定
④ EMT誘導なし及びありの三次元遊走モデルによるEMT誘導細胞の
遊走の定量化
⑤
FN
gradientを組み込んだ三次元遊走モデルによるEMT誘導細胞の遊
走の定量化
⑥ α5-integrin阻害剤であるRGDSを用いた際の三次元遊走モデルによ
るEMT誘導HPMCの遊走の定量化
⑦三次元遊走モデル条件における細胞増殖試験
⑧ EMT誘導fIPMCにおいてFN添加およびRGDS阻害時のα-SMAの
発現量の測定(ウェスタンプロット)
3-1.実験系の作成
ヒト腹腔内体網から採取した腹膜中皮細胞を培養して、以後の実験を行っ
た。ヒト体網からの腹膜中皮細胞の採取(初代培養)及び継代培養は生体材
料化学研究室の共通プロトコル(APPENDIXA-1.細胞培養プロトコル参照)
に従った。ここではその概略を記す。
3-1-1.ヒト大網由来腹膜中皮細胞(EPMC)の培養
本研究で使用したHPMCは、予め本学医学部偉理委員会によって患者組
織の採取につき承認を得て、
informed
consentを得た患者からいただいた体
網組織に由来する。腹腔内手術から得られたヒト体網塊を親指大の切片に
切り分けた.
10%trypsinnBS溶液により
HPMCを単離・採取し、各種
supplement及び10%FBSを添加したM199を用いて培養フラスコ内で培養
した。本研究では継代数が2-3代目の細胞から細胞密度が5×105個/mlの
細胞懸濁液を作成し、
6穴プレート及び¢35シャーレ、
24穴プレートに播
15
種した後、
connuentまたはsubconf)uentまで培養したものを使用したD培地
は3日に1回交換したo
3-l12.
ⅡPMC
-のTGF-β
1誘導
6穴プレート及びd'35シャーレにsubconnuentまで培養したHpMCの培
地を、
o.1%FBS/M199と交換し24h
12、
TGF-β1/M199と交換し、
3-2.
TGF-β
growth
24、
arrestを行ったoその後10ng/ml
48h培養した。
1誘導による細胞形態変化の観察(動画撮影及び静止
画撮影)
動画撮影
¢35シャーレに培養したHPMCにTGF-β1を誘導後、倒立型位相差顕
Tokyo,
微鏡IX50(OLYMPUS,
Japan)にセットし(Fig.1-I)、取り付けたMOS
により画像をx-capにて保存したo画像取り込みを1枚/10分行い、
72hま
で行ったo保存した画像を画像処理ソフトVirtualDub(Vcr.1.617著作Avery
Lee氏)を利用して動画作成を行ったo
細胞の入ったシャーレを入れる
Fig.
3・1シャーレの設置場所
r
l
】
[
I/:Lr
i
16
静止画撮影
24、
TGF-β1誘導後,各時間(o、
48b)におけるHPMCの形態を位相差顕
微鏡cK2にて観察し、デジタルカメラCAMEDIA
C-3040
ZOOMにてその画
像を撮影し、パーソナルコンピューターに取り込んだ。
3-3.
l誘導によるSnai1及びE-cadherin、
TGF-β
MMP-2
mRNAの経時的測定
作成したHPMCのTGF-β1誘導モデルより、
E-cadherin、
α
Snail及び
AGPC
mRNA発現量の経時的測定を行った.
RT-PCR法,電気泳動法は生体材料化学研究室の共通プ
ロトコル(APPENDIXA-3.
A-4.
mRNAの抽出し、
5-integrin、 MMP-2
法、 NAの定量法、
ル、
α5-integrin、
AGPC法プロトコル、
A-3.
RNAの定量プロトコ
RT-PCRプロトコル参照)に従った.ここでは,簡単にその概要を
記す。
3-3-I. Acid
Guanidium-Phenol-Chloroform法(AGPC法)によるmRNAの
抽出と定量
作成したHPMCのTGF-β
1刺激モデルにRNA-BeeTM
RNAを1ml/well加え細胞を掻き取った。
chlorofbm
-
OF
ISOLÅrION
isoamylalcobo1
24:lを
200〟1加え15分間静置した(室温)0
12000G/4℃/15分にて遠心分離,最上
層より350〃1採取し、
isopropano1を500JLl加え-80℃にて一晩保存した.
解凍後12000G/4℃/10分にて遠心分離、上澄を除去し、
理水溶液を500FL l加えたo
75%EtO矧DEPC処
10000G/4℃/5分にて遠心分離、再び上澄を除去
17
し、減圧乾燥後DEpC処理水を10〃
Smart
SpecTM
3000
1加え、55℃/5分加温後4℃に冷却した。
Spectrophotometerを用いて各サンプルのmRNAを定量し
た。
3-3-2.
mRNAの逆転写によるcDNA作成とpolymerase
(PCR)によるSnai1及びE-cadherin,
α5-integrin
Reaction
Chain
cDNAの増幅
mRNAの定量結果を基に、全サンプルのmRNA濃度を統一した。これら
ScriptTM
のmRNAをSuper
RNase
H
Reverse
Transcriptase及びRandom
primersを用いてcDNAに逆転写した(RT
product 2
productの作成).得られたRT
GoldTM DNApolymerase
JL lと、各種primer(Table.3-1)及びAmpliTaq
を含むPCRMasterMixを48JLl混合し、サンプル内のcDNAをpCRにより
増幅させた(PCR
productの作成)。等量のcDNAが用いられているかを確
認し、 PCRproductを半定量化するため、内部標準としてbouse
keeping
gene
であるGApDⅢを用いた。
Table.3-1
PrimerList
Size
Sequence
Primer
Cycles
O)p)
5'-GATGTCATCATAmGGCAGGTT-3'
Sense
GAPDH
29
322
38
487
36
500
39
453
29
292
5'-CCTGCACCACCAACTGCTTAGCCC-3'
antlSenSe
5'-TCAGACGAGGACAGTGGGAAAG-3'
Sense
Snai1
5'-GCTTGTGGAGCAGGGACATTC-3'
aJ1tiSenSC
Sense
5'-TCCCÅrCAGCrGCCCAGAAA-3'
antlSenSe
5'-TGACTCCTGTGTTCCTGTTÅ-3'
E-cadherin
5'-GGCATCAGAGGTGGCTGGAGGCIT-3'
Sense
α5-integrln5'-GGCAGCTÅrGGCGTCCCACTGTGG-3'
antiscnse
5'-ACAGCCTGGCÅrGGGGCAAGT-3'
Sense
MMP-2
5'-TTCTCCTCCÅrCCAGTGGAG-3'
antlSenSe
3-SIS.電気泳動とバンドの定量
wtルol Agarose
pcRによる増幅後、3%
を含む600mlの1x
Tris-acetate-EDTA
の条件でPCRprodud
gelと、0.3
FL
g/mlのethidium
buffer泳動液を用い、
bromide
80V/80mA/70分
lO〃1の電気泳動を行った。泳動後、イメージアナラ
イザーにてjpeg型式でゲルの画像を撮影し、画像解析ソフトGe1-Pro⑧
Analyzer
Version
3.1 for
WindowsTMを用いてバンドの輝度を定量化した.結
果はGAPDHの値に対するSnail及びE-cadherin、
三重大学大学院
工学研究科
α
51integrinの値の比とし
18
て示した。
3-4.化学走化性因子を用いたEMT細胞の三次元細胞遊走評価
具体的な方法はAPPENDIX.
に従
三次元細胞遊走モデルの作製
A-6
った。ここでは簡単な方法を示す。
3141l. EMT誘導なし及びEMT誘導における三次元細胞遊走評価モデルの
作成
24穴プレートに10pg/mlFNを含んだ1.5mg/mlcollagengelを200FLl加え、
37℃/30分インキュベ-トしてゲル化させた(controlとして1.5mg/ml
0.5mg/ml
collagengelを用いた)。さらにそのゲルの上に、
collagengelを200
FNの拡
37℃/30分インキエペ-卜してゲル化させることにより、
111加え、
散層を作成したo FNは48h、
72hインキュベ-卜して濃度勾配を作成した.
またゲル上に膜孔121Lmのインサートを乗せ、
0.1%FBS伽199
誘導条件として10ng/ml
EMT
で、また
EMT誘導なしでは
TGF-β
l加えた(Fig.3-3a)c
1/0.1%FBS/M199を300FL
予め¢35シャーレにsubconnuentまで培養し、
Ⅰ廿MCに10ng/mlTGF-β
EMT誘導条件のときのみ
1を48h曝露したoそれを継代操作し,細胞数が5
×104個/穴となるように作成しておいた24穴プレートのインサート上部播種後、 72b培養した(Fig.3-3b、
Fig.3-3c)0
T
㌔*/地′
l
)?n(・
・
:
F
J′
i
L事7
ロト\\・・蒜・I
L:::.
・14・.P
col甲ellgeJ
`
L▲
■.■
L+-
T⊥
⊇
-血」転」一-
Lb・〒_II__b
a
i.-..t・
-
C
Fig.3-3三次元細胞遊走評価モデル
3-412.
FN及びBSAを用いた三次元細胞遊走評価モデルの作製
31411の実験と同じように24穴プレートに、
1,5FLg/ml
BSAを含んだ
1.5mgmi
collagen
gelを200FLl加え、
また同じように上層に0.5mg/ml
37℃/30分インキエペ-卜してゲル化させた。
1加え, 37℃/30分イン
co11agengelを200〟
キュベ-トしてゲル化させることにより、
FN及びBSAの拡散層を作成し
た。
FNの濃度勾配と一致させるために,
BSAの拡散時間はそれぞれ16h、
24bインキュべ-卜してそれぞれの濃度勾配を作成した。
遊走実験に使用した細胞はEMT誘導条件の細胞を用い3-5-1と同条件で
19
実験に用いた。
3-4-3.
RGDS阻害剤によるFN存在下での実験条件
FNのゲル内の濃度及び拡散時間等は3-5-1に記した.
RGDSによる阻害実験では、まず予め¢35シャーレにsubcon且uentまで
に培養した細胞に10ng/ml
TGF-β
1を48h曝露した。それを継代操作する
際に、 5JLg/mlのRGDSを曝露し、
30minインキュベ-トを行った。その後
継代培養するときに、細胞数が5×104個/穴となるように作成しておいた
72h培養した。
24穴プレートのインサート上部-播種後、
3-4-4.遊走細胞数の計測
以上の実験条件で行った三次元遊走細胞の測定方法について以下に記す。
まず三次元遊走実験に用いた培地を取り出し、そこにPBSをいれ、
PBS
を取り除いた培地と混ぜ、上澄みとした。その後インサートを取り除き、
インサートは風乾し、ディフクイック染色(ApPENDIX参照)により細胞数
を測定した。また上澄みは遠心分離して溶液を取り除き、
M199の50〃1程
度細胞懸濁液を作成した。血球計算盤にて細胞数をカウントし、細胞数を
計算した。
ゲルにcollagenase
S-1を30〃1加え、
37℃/30分インキュベ-卜してゲル
を分解した。その溶液を遠心分離して上澄み液を取り除き、
M199を加えて
50〃1程度の細胞懸濁液を作成した。血球計算版にて細胞数をカウントし、
細胞数を計算した。
3-4-5.細胞増殖試験
具体的な方法はAPPENDIX.
A-7
細胞増殖試験
に従った。ここでは
簡単な方法を示す。
10%FBS/M199で24hイン
¢35シャーレにHpMCを5×104個播種した。
キュベ-トし、細胞を接着させた。その後o.1%FBS/M199で24hgrowha汀eSt
行い、各シャーレごとに以下に示す条件で培養を行った。
●
10%FBS/M199
●
0.1%FBS/M199
●
0.1%FBS刀M199+
long/mlTGF-β
1
●
0.1%FBS/M199+
long/mlTGF-β
1
+
10Jlg/mlFN
培養24bごとに96bまで各シャーレの同一箇所を写真撮影し、細胞数を
カウントした。
三重大学大学院
工学研究科
20
3-5.三次元遊走実験条件下でのα-SMAのウェスタンプロット測
定
三次元遊走モデル条件下で培養したHPMCより、
●
0.1%FBSバM199
●
o.1%FBS/M199+
long/ml TGF-β
1
(24h曝露)
●
o.1%FBS/M199+
long/mlTGF-β
1
(24h曝露)
1
+
⇒10
●
p
g/ml
FN(24h曝露)
0.1%FBS/M199+
⇒5
p
long/mlTGF-β
10pg/mlFN
g懲G%D芸冨口.'.3:m3m:FN72:ご蒜
タンパク質を抽出し、
α-SMA発現量の経時的測定を行った。タンパク質
SDS-PAGE、ウェスタンブロ
の抽出法及びBCA法によるタンパク質の定量、
ッティング、抗体反応とバンドの定量は体材料化学研究室の共通プロトコル
A-6
(APPENDIX.
参照)に従った。ここでは、簡
ウェスタンプロット法
単にその概要を記す。
SIS-I.タンパク質の抽出とBCA法による定量
上記の実験条件の細胞に、細胞溶解液を300〃1加え、さらにprotease
inbibitor及びpbosphatase
inhibitorをそれぞれ2〃
1ずつ加えた。室温で15分
ほど軽く浸塗した後、セルスクレーパーで細胞を掻き取った。冷却遠心機
で15000rpnJ30分遠心し、上澄み液を取ったo抽出したタンパク質溶液は
-
20度で冷凍保存した。
A溶液とB溶液を50:1で混合したもの100〃1に対して、サンプル及びス
タンダード(BSA)を10plずつ加えた。
37℃/30分静置し、
OD562を測定
した。スタンダードのタンパク質濃度を横軸、吸光度を縦軸にとり、検量
線を作成した。その検量線を元にサンプルの総タンパク質濃度を測定した。
3-5-2.
SDS-PAGEによるタンパク質の分離
ゲル板にruming gel (10%アクリルアミドゲル)とstacking
gel
(4.5%ア
クリルアミドゲル)溶液を加え、コ-ムを取り付けて30分間静置してゲル
21
化させた。コ-ムを取り外し、ウェルを作成した。泳動バッファーを満た
した電気泳動槽にゲル板を固定した。抽出したタンパク質溶液とサンプル
バッファーを1:1で混合し、熱湯100℃/3分でボイルした。各ウェル内にサ
15mA/2
ンプルを40〃1程度及び分子マーカーを5〃1ゆっくりと注入した0
時間泳動を行った。
3-5-3.膜-の転写(ウェスタンブロツティング)
電気泳動終了前にmming
gelと同じ大きさに切ったPVDF膜をメタノー
ルに20秒浸し、さらにB溶液に30分浸塗させた。またmnninggelと同じ
大きさに切ったブロツティング漉紙をA溶液に2枚、
B溶液に1枚、
c溶
液に3枚浸した。電気泳動終了後、running
gelをB溶液に浸した。ブロツ
ティング装置に、準備していたA溶液に浸していたプロツティング洩紙2
枚を乗せ、さらにB溶液に浸していた漉紙1枚、
C
pvDF膜、runninggel、
溶液に浸していた漉紙3枚の順番に上に重ねてセットした。
72mA/40分ブ
ロツティングした。ブロツティング後pvDF膜をブロッキング液に浸し、
overnigbtブロッキングした。
3-5-4.抗体反応とバンドの定量
α-sMA一次抗体溶液をpvDF膜に添加して、室温下で1時間静置した。
その後pBSTで10分毎に3回洗浄した。洗浄後二次抗体をpvDF膜に添加
して室温下で1時間静置した。再びpBSTで10分毎に3回洗浄した。
pvDF膜上に添加し、
ECLl液と2液をそれぞれ750〃1ずつ混ぜ、
静置した。 Typboonでバンドをスキャンし画像を保存した。保存した画像
をImage
1分間
quantで定量を行った。
:._求)( I?: )( I?I:ド';i:f'_ノi・[:研'先手こf.
22
4.結果
4-I.
TGF-β
1誘導による細胞の形態変化の観秦
TGF- β1誘導したときの、細胞形
subconfluentに培養したHPMCに10ng/ml
態の経時変化を、デジタルカメラにより撮影した画像を示す(Fig.4-1)。
TGF-
β 1誘導直後(oh)では細胞間接着がしっかりしており、敷石状の形態であっ
たo TGF-β
1の誘導時間の経過にしたがって、細胞間が離れ形態が紡錘状に
細長く変化した。
Oh
Scale: 250トLlll
4Sh
FigAll
TGF-Pl誘導によるHPMCの形態変化(scale:250岬l)
23
4-2.
TGF-β
1誘導による細胞の巷走距離の測定
Subconnuentに培養したfIPMCをo.
long/mlTGF-β
1%FBS/M199で24h
growth
arrestを行い、
1曝露し、 10分毎に撮影を行ったo撮影した写真をAPPENDⅨ
で記したImageJを使い,細胞の遊走距離をIh毎に測定し、
6h毎に平均を取
った。 TGF-βlの曝露によって4-1で記したように、細胞の形態変化が見ら
れ、曝露後24-36hで遁走距離が最大となった(A-20)OTig.4-2)D
(
≡
580
E∃
讃60
塁40
愚
6
12
18
24
30
36
42
48
54
60
66
72
TGトβ 1曝露時間(hr)
Fig.4-2 TGF-β
1誘導によるⅠ廿MCの遊走距離
](i::ノJ-l'r:iLIE
・T・=-
L羊研']Lf列
24
4・3.
TGF-β
MMP12
1誘導によるSnail及びEICadherin、
a5-i血tegrin、
mRNAの経時的珊定
以下に示すsnai1及びE-cadherin、
α
5-integrin、 MMP-2
mRNA発現結果に用
いた値は, RT-PCR法によって激定したそれぞれの発現量を、内部標準である
GAPDHmRNAの発現量で除し、
4-311.
controlが1となるように補正を行ったo
TGF-
β1誘導によるSnaiJ mRNAの経時的測定
Fig.4-3にTGFβ1 (10ngml)誘導に対するSnaiJ皿RNA発現の時間変化
を示す。
TGF-β1の誘導後からSnai1
mRNA発現は増加した。また誘導24
48hではやや減少
controlの約1.8倍の発現を示した。
時間後で最大となり,
したが、
controlよりも高い発現レベルを維持していた(n=S)。
Snail
GAPDH
TGFヰ1誘導時間
Oh
:
12h
p旬.00
24h
48h
‡
1
O
b
⊂
○
く)
u)
>
=
⊂l
EL
<
(コ
g≡一
喜
¢ontro
Fig.4-3
1
1 2h
TGFIPl誘導によるSmil
24h
48h
lllRNAの経時変化
25
4-3-2,
TGF-
βl誘導によるEICadherin
mRNAの経時的測定
β 1 (long/mi)誘導に対するE-cadherin
Fig.4-4にTGF-
mRNA発現の時間
変化を示す。誘導に対してE-cadherin
mRNAは時間依存的に減少したQ
た誘導48時間後ではcontrolの約o.7倍の発現の減少を示した(A-8)o
ま
E-cadherin
GAPDH
TGF-β1誘導時間
Oh
12h
48h
24h
O
L.
■lJ
⊂
O
O
の
>
=
(⊃
CL
<
O
0.8
0.6
i?
t=
-⊂
0.4
4)
」=
-1⊃
匂
O
0.2
I
U」
Oh
Fig.4-4
12h
TGF-β1誘導によるE-cadherin
:.
・Ti;人
Tr=
48h
24h
i(;I:∃完
I
mRNAの経時変化
i;:卿'JjL-H:.ト
26
4-3-3.
TGF-
β 1誘導によるα5-integriJI mRNAの経時的測定
FigA15にTGF-β
1
(long/ml)誘導に対するα
5-integrin mRNA発現の時間
変化を示すo
TGF-β l誘導に対してα5-integrin mRNAは12hの時点から強
い発現を示し、
48bでもその強い発現を維持していた(n==12)0
a
5-integnn
GAPDH
12h
TGF-β1誘導時間oh
24b
48b
=o
.トI
S
く)
くゎ
>
=
⊂l
EL
a
⊂
■⊂
hD
4)
lP
⊂:
-a;
es
¢on廿o1
Fig.4-5
1 2h
24h
48h
TGF-Pl誘導によるcL5-integinmRNAの経時変化
27
4-3-4.
β1誘導によるMMP12
TGF-
1
Fig.4-6にTGF-β
化を示す。
を示し、
TGF-β
mmAの経時的軸定
(long/ml)誘導に対するMMP-2
mRNA発現の時間変
1誘導に対してMMP-2
mRNAは12bの時点から強い発現
48hでもその強い発現を維持していた(n-6)0
ML4P-2
GAPDH
TGF-β1誘導時間
12h
oh
24h
4Sh
O
」
■J
】=
O
O
(○
>
=
⊂】
%
O
6ヨ
5コ
I
A.
≡
≡
co
fig.4-6
12h
ntrol
24h
TGF-β1誘導によるMMP・2
:.
)(']y 】▲,L
LTl:)( pL3・':
48h
mRNAの経時変化
l
折''j4.E
[=:】
28
4-4.化学走化催因子を用いたEMT細胞の三次元細胞遭走評価
4-4-1.
FNを用いた遊走細胞数の計測
FNの
Fig.4-7に三次元遊走モデルを用い,化学走化性因子として下層濃度勾配を形成し、膜下及びコラーゲンゲル中-遊走した細胞を測定し、
FNなしのときの遊走細胞数を1として結果を示すc
態に比べて、
FNを含んでいない状
FN濃度勾配が存在すると下層-遊走した細胞数は増加したo
下層に遊走した細胞数はFNの拡散48hでControlの1.6倍になり、
2.1倍もの細胞が遊走した。
72hで
(a-5)。
O
a
⊂
O
I)
軍1.5
萱.
層o.5
0
w仙out
FN
diffusion
diffusion
48h
Fjg-417下層-遁走した細胞数vs
4-4-2.
72h
COJ)trO]
BSAを用いた遁走細胞数の計測
Fig.4-8に三次元遊走モデルを用い、化学走化性因子として下層度勾配を形成し、膜裏及びコラーゲンゲル中-遊走した細胞を&r)定、
しのときの細胞数を1としたグラフを示すo
て、
BSAの濃
BSAな
BSAを含んでいない状態に比べ
BSA濃度勾配が存在すると下層-遁走した細胞数は16hで1.2倍に、
24h
で1.4倍の細胞数-とわずかに増加したが、有意差は見られなかった(n-5)。
○
1.6
ヽ_
盲
O
O
1.2
∽
>
き謂0.8
響
罪
0.4
without
BSA
difhsion
16h
Fig.4-8下層-遊走した細胞数vs
diffusion
24h
control
29
4-4-3.
FNを用い、
RGDSにより阻害した際の遊走細胞数の計測
Fig.4-9に三次元遊走モデルを用い、
EMT誘導しなかったもの及び誘導し,
integrinの阻害剤RGDSを添加したときの結果を示す。これは化学走化性因子
として下層-
FNの濃度勾配を形成し、膜下及びコラーゲンゲル中-遊走し
た細胞を測定した結果で、
EMT誘導ありFNなしのときの遊走細胞数を1と
したもののグラフである。
EMT誘導していないものは、
比べ遊走していないことが分かるoまたRGDSによる阻害は、
EMT誘導したものと
EMT誘導細胞
の遊走をかなり抑えることが分かった(n-3)0
ト-pg.o41---」
TGF-β
1
+
FN
RGDS
Fig.4-9下層-遊走した細胞数vsEMT誘導HPMC
I
市-/、こ′-)i.'-Lノ、rLi::院
j;:朋'Jit T;.l
30
4-4-4.細胞増殖試敦
Fig.4-lOに各条件下において細胞増殖-の影響を測定した結果を示すo条
件は以下の4種類で行った(a-3)。
●
10%FBS/M199
●
0.1%FBS/M199
●
0.1%FBSノM199+
long/mlTGF-β
1
●
0.1%FBS/M199+
long/ml
1
TGF-β
+
10〟g/血IFN
通常の細胞培養条件下である10%FBS/M199では細胞は増殖し続けたo
方gfOWth
一
arrestやEMTを起こさせる条件化では細胞は増殖しなかった8
2
1.8
1.6
云
7.4
∽
1.2
>
a
餐
暴
1
0.8
0.6
0.4
0.2
0
1 0%FBS
0.1 %FBS
TQF-
a
TGF+
FN
Fig.4-10細胞増殖の割合
31
4-4.
TGF-β
1誘導によるα-SMAの轟時的軸定
Fig.4-11にFNがα5-integrinに接着した際のα-SMA発現の時間変化を、ウ
ェスタンプロット法により測定した結果を示す。実験条件は4種類作り、以
下に記すo値はα-SMAの発現量においてcontrolが1となるように補正を行
ったo control以外では非常に顕著な発現が見られた(n=3)。とくにTGF-β1
曝露後、 FNを添加したものはα-SMAの発現が大きく見えたo
このことは
RGDSで阻害を行うことでTGF-β
1曝露と同じ程度の発現になることからも
伺える。
実験条件
①o.1%FBS/M1
99
②o.1%FBS/M199
+
long/ml
TGF-
β1(24h曝露)
③o.1%FBS/M199
+
long/ml
TGF-
β 1 (24b曝露)
TGF-
β 1 (24h曝露)
⇒
10JLg/mlFN(24h曝露)
+ long/ml
④o.1%FBSnu199
⇒
RGDS阻害剤添加(30分インキュべ-ト)
⇒10FLg/miFN(24h曝露)
①
a-
S加IA
「6
b
5
8・00
⊂〉
【○
>
6.OO
・<
≡
u)
1
4.00
eI
TGF-β
1
FN
RGDS
Fig.4-11三次元遊走実験条件下でのα
垂)、二号)、こrt-i
I:I;i
r
-SMAcontrol
;I:G]r.雅章:】
32
5.考察
1の産生機序
5-1.腹膜透析におけるTGF-β
高濃度glucoseやグルコース分解産物(GDP)によってHPMCでは、
PKC
(proteinkinase C)の活性化が起こる【35]。活性化PKCにより脱リン酸化した
転写因子c-Fosとc-Junの複合体(AP-1
protein-1)は、 TGF-β 1遺
伝子上のプロモーター領域のAP-1結合部位に結合し、TGF-β
1
mRNAが発現
:
activated
【16]o
する.それによりTGF-β1の産生が促進される(Fig.511)
路によって、
HPMCではTGF-β1がオートクリンに産生され、
このような経
TGF-β1が
HPMCに作用する事でEMTが進行すると考えられる【8]。
細胞膜
′
′
J
∫
I
I
l
I
l
∼
ヽ
ヽ
I
TGF-BI
■■
/
▲l
■■
■■
_
Fig.5-1
TGF-Plの産生経路
33
5-2.
1によるSnai1
TGF-β
mRNAの
mRNA発現に伴うE-cadherin
減少メカニズムとその役割
本研究においてTGF-β1をHpMCに作用させたとき、
TGF-β
Snail
mRNAは
1誘導12時間後に有意な発現の増加を示したのに対して(Fig.4-3)、
24hで有意な減少を
mRNAは12時間では有意な減少は見られず、
示した(Fig.4-4)。このようなmRNAは以下のシグナル伝達経路を経由するこ
E-cadherin
とが考えられている。
TGF-β
1がHPMC細胞の膜上に存在するTGF-β1レセプター(セリン/ス
レオニンキナ-ゼレセプター)に結合すると、レセプターの細胞内ドメイン
にあるセリン/スレオニン残基がリン酸化する。続いてTGF-β1が結合したレ
セプターがエンドサイト-シスし、
(ホスフ
SH2/SH3ドメインを有するPI3K
PI3Kは細胞
ァチジルイノシトール3-キナ-ゼ)がリン酸化部位に結合する。
膜上のPIP2
(ホスファチジルイノシトール4,5-ビスリン酸)をpIP3
ァチジル3,4,5-トリスリン酸)
for Receptor
(ホスフ
(SmadAncbor
-とリン酸化する。続いてSARA
Smad2/3は
Activation)はPIP3及びTGFレセプターと結合する.
sARAを介してTGFレセプターと結合しリン酸化される【36,37】。リン酸化した
smad2/3はCo-SmadであるSmad4と複合体を作ることで核内-と移行し、
snail
mRNAのプロモーター領域に作用する事でSnail
mRNAを発現させる
【11,36]。またもう一つの経路としてMAPKカスケードがある.それはRas、Raf,
MEK、
ERKという経路のリン酸化を通して活性化し、
せる【36]o
これらの経路によって発現したSnail
プロモーター領域に作用する事で、
Snail
mRNAを発現さ
mRNAの
mRNAはE-cadherin
E-cadberin
mRNAの発現を抑制する。
adberence junctionを構成するE-cadberinは細胞間接着を維持する事で、細胞
の形態を維持している。しかしTGF-β
1などによってE-cadberinが減少する
と、細胞間接着が失われることで形態変化を起こし、間英系細胞のように遊
走能を持つものと思われる。また細胞間接着の指標(EMTの指標)としては
tight
junctionの構成成分であるZO-1やoccludinsなども用いられている【3,14】。
zo-1やOccludinsはglucose添加によってリン酸化され、
質透過性の克進に関与する主要なタンパク質である【38].
tight
down-regulateし、溶
adherence
junctionや
junctionはそれぞれが細胞内で細胞骨格タンパク質アクチンと結合して
いることから、
EMTと溶質透過性克進の間には関連性があると思われる。
:.車l:′羊人J、i::院
「ノ、i::研究村
34
Fig,5-2
TGF-β1によるSnai1 mRNA発現メカニズム
これらのシグナル伝達メカニズムから、
snail
mRNA発現に遅れて
E_cadhe血mRNA発現の減少が起こると考えられる。
35
5-3.
MMP-2
EMT誘導ⅢpMCにおけるα5-integrin,
mRNAおよ
びα-SMAの発現と遊走における役割
細胞の遊走に大きな役割を果たしていると考えられている因子すなわち、
5-integrin、
α
MMP-2、
1によるHpMCからの発現を調
α一SMAに着目しTGF-β
べた.これらの因子について簡単に記す(Table.5-1)
Table.5-1細胞遊走因子とその役割
問質層-の遊走における役割
FNと結合することで様々な遊走に
関するシグナルを細胞に伝達する
α5-integrln
MMp-2
問質層中のCo11agenを分解する
α-SMA
細胞遊走に必要なストレスファイバー
このような役割を持った因子は、
TGF-β
1誘導によってα5-integrin
mRNA
の強発現(Fig.4-5)、MMP-2 mRNAの強発現(Fig.4-6)が見られ、さらにはα
の強発現が見られた(Fig.4-ll).また3次元遊走実験において、EMT誘導I廿MC
と誘導なしのHPMCの遊走細胞数を比較すると、
-SMA
EMT
EMT誘導なしに比べ、
誘導のほうが遊走細胞数が多かった。この遊走細胞数の増加は、
導により増加したMMp-2が下層のcollagenを分解し、
TGF-β
1誘
α-SMAの発現により
遊走能が高まったためだと考えられる。
このMMP-2やα5-integrinのTGF-β
1誘導による発現はSnail
mRNAと同じ
シグナル伝達経路で発現すると考えられている(Fig.5-2)。一方α-sMAの発現
は、まずTGF-β
1が細胞膜上に存在するTGF-β
1レセプターと作用する事で、
低分子量Gタンパク質のRhoファミリーであるRhoAを活性化することで始
(mammalian
まる。この活性化したRhoAはmDia
diaphanous)、
DIP
(diaphanous-interacting
protein) Arp2/3複合体を経由またはmDia、プロフィ
リンを経由してα-SMA発現を増加させる。一方でRboAはRboキナ-ゼで
、
あるROCK
(Rho associated coiled-coil-containing protein kinase)と結合し、活
性化ROCKはMLC
(myosin light chain)をリン酸化してα-SMAの会合を促
進する(Fig.5-3)oこれらはRhoAやROCKの阻害実験により実証されている
[14,39]
0
:.蚕大'?-'人Jl;I:院
T-.'、芦研究科
36
細胞膜
RhoA
15i
iiZF
ROCK
mDi&
/,
プロフィリン
DIP
1
@i@
Arp2/3複合体
iiZ
α-SMA
I
iiI
/
細胞骨格再構築
Figj-3
TGF-β1誘導によるα-SMA再構築のシグナル伝達経路
このようにTGF-β
1誘導を起こしたHpMCは,
MMP-2やα-SMAを発現す
ることで遊走能が増加すると考えられる。
さらにα-SMAの発現に注目すると、TGF-β
1誘導HPMCにFNを添加した
とき、 TGF-β1誘導のみのものに比べ、発現の増加が見られたPg.4-ll).こ
のことからTGF-β1誘導HPMCにFNが作用するときさらに遊走細胞数が増
加することが考えられる.実際三次元遊走実験において、下層のco11agenゲ
ル中にFNの濃度勾配を形成したときに遊走する細胞数が、下層にFNを含ま
ないものに比べ増加した伊ig.4-7)o一方下層にBSAの濃度勾配を形成したと
きは,
EMT誘導HPMCの遊走細胞数は、下層に何も仕込まなかったもの
(control)と有意差は見られなか<TDた伊ig.4-8).さらにFNの接着を阻害する合
成ペプチドRGDS
を添加すると、下層にFNの濃度勾配を形成させ
EMT(+)HPMCの遊走細胞数は、
EMT(I)HPMCと同程度に遊走細胞数が減少し
た(Fig.4-9)oこのようにFNの添加により遊走細胞数が増加する原因は、
TGFβ 1誘導によって強発現したα5-integrinがFNに結合したためと考えられる
咋ig.4-6).
以下TGF-β
1誘導α5-integrinの発現とFNとの結合について考察すると、
前記したようにSnailの発現と同じシグナル伝達経路によって発現すると考
えられている(Fig.5-2).図5-4に示すように発現したα5-integrinはFNと結合
することで、 FAR(Focal adhesionkinase)をまずリン酸化する. FAKのリン酸化
に伴いSrc(非受容型チロシンキナ-ゼ)の活性化が引き起こされ、同時に
Paxillin(接着斑構成タンパク質)においても活性化がおこる。これら2つの経
路の内、まずsrcを経由するシグナル伝達経路では、
ERE(Extracellular
37
KlnaSe)を通り、接着斑のターンオーバーを行う経路とp-130
signal-Regulated
Cas
(Crk associated
substrate)やRac(Small GTPase), JNK (Jun amino-terminal
kinase)を経由し、 MMP-2(コラーゲン分解酵素)を産生し、問質層-の遊走を促
進する経路にシグナルが平行して伝わるoまたPaxillinを経由する経路も平行
して活性化し、
Racを経由し接着斑のターンオーバーや葉状仮足の形成、ま
たさらにATP2/3
(Actin-relatedprotein)を経由しα-SMAの形成に働く【5S】.
HeparlnⅡ
blndln9
q5Bl-htegrln
9(to
bhldln9
I
HepaTln
SIte
&Ⅱ
blndh19
SJte
Src
、qiEI
■
ER
K
p
I
130竿-.■
foTTT)atJorlJ_I
_.(I--㌫&Ye7
ー
,A,,2n
\
c且IT
Foe.1l.1dLheヽlcn
nn?TV}L一
⊥-㌔-・-_千
alrl
JNfK
十
MhTP-2 mRNA
ー(
_--
/LaTTnlJ
Ipod(8
-
■1-I..■rーl'"I
lormatlon
--・・t細胞核
l
-
-
ーーーーー」
-㌔
(: Ce[lhva91on
I
''
--二rT二rrご(-
このように、
-一一--
Fig.5-4
integrin-FN接着によるcL-SMA再構築のシグナル伝達経路
TGF-β1により強発現したα5-integrinはFNと結合し、
Talin
といった接着斑のターンオーバーやMMP-2
胞の問質層-の遊走(Gen
mRNAを発現することによる細
hvasion)の促進や,葉状仮足の形成,
α
-SMA(Stress-h)の形成などを引き起こすことから、や5-imiがFNと結合
一il:.:・
!j
ILt主IililJL=・Li
u'iiiii;ニ辿土ユ立上エ
⊥エIi:I
Ji i・二・1l・1二⊥二L辿_三__」⊥11J.
≡.
_:
イトケラチン什)の線維芽様細胞の源が腹膜申皮細胞であることを裏付ける重
となった
T,.:セノj::人:J:l・こ';E [
]・・
ET]r'先手:i
38
5-4.
FN阻害剤としてのRGDSの妥当性
EMT誘導HPMCとFNの結合が細胞遊走の実態であることを、さらに明確
にするため、両者の結合阻害剤RGDS[59](Arginine-Glycine-Aspartic
acid-Serine)
アミノ酸配列RGDSは、緒言に記したようにFNに含ま
を用いて検討したo
れるintegrin接着部位である(Fig.1-ll)Q
これをFN添加前に30分インキュベ
FN-integrinの結合の阻害という実験を行ったo三次元遊走
-卜することで、
実験において、
RGDSによる阻害を行ったEMT誘導HPMCは、下層にFNが
存在する条件においても、ゲル中や膜下に遊走する細胞数が、
RGDSによる
阻害を行っていないものに比べると遊走細胞数が顕著な減少が見られたこと
からRGDSによる阻害が十分に行われていることを示している(Fig.4-9).ま
た三次元遊走条件下でのウェスタンプロットにおいて、
RGDSによる阻害に
よって、
α-SMAの発現が一部減少したことからもRGDSによる阻害ができ
ると考えられるO;ig.4-ll)o
しかしRGDSをEMT誘導HPMCに作用させるだけでは、
integrinが接着し
たことと変わらないのではないかという疑問が生ずるが、これはFN
と
integrinの結合機構とシグナル伝達を詳しく考察することで解決できる.まず
integrinがFNのRGDループ畔GDS)に強く結合し、FNとのアンカーの役割を
果たすoその後,
integdnに近接しているSyndecan(HSPG
‥ Condroitin
:
Heparansulphate
surface
proteoglycans)L53][54】が-パリン結合
部位OTig.1-9及びFig.1112)に作用することで、
integrin下に接着しているFAK
proteoglycan及びCSPG
を活性化,その後Fig.5-4の経路やRhoA
(RA各 homolog
gene
family, member
A)
を経由することでα-SMAを形成するようになるO;ig.5-5)
Hepal]nE
Fig.5-5
blndhg
s紬
45Bl・Lnteg†lhblrdLng91t●
H叩arJnI
&ZL
bJ11dTrI98tfe
Syndecanとintegrinの相互作用によるシグナル経路図
39
このシグナル伝達経路図にあるように、
FNが細胞に作用するとき、
Syndecanとintegrinが非常に近い位置にあり、同時に作用しあわなければ活性
化しない。本実験のように、
RGDSによりあらかじめ細胞の血egrinの接着を
行ってしまうと、 SyndecanがFNのHeparin結合部位に接着することができな
くなり,それによりシグナル伝達ができなくなることから、
RGDSによる阻
害ができると考えられる。
5-5.総括
本研究ではコラーゲンゲルを積層させることによって化学走化性因子の濃
度勾配を任意に形成し、下層-遊走した細胞を定量的に測定するという新規
三次元遊走モデルを作成した。このモデルは今までの遊走モデルにはなかっ
た炎症時に形成されるであろう問質中の走化性因子の濃度勾配を考慮したも
ので、より生体に近い遊走モデルである。
このモデルを使うことで、
EMT誘導HPMCの問質層-の遊走にFNが関与
している可能性が示された。これにより、腹膜透析における腹膜線維症や腹
膜劣化抑制など、腹膜透析の更なる長期継続を可能にできるような材料開発
を可能とする。さらにはこの遊走モデルは腹膜組織のEMT誘導HPMCの遊
走メカニズムの解明だけでなく、ガン細胞の浸潤メカニズムや器官形成時の
細胞遊走メカニズムの解明などさまざまな分野-の応用が考えられる。
三重大学大学院
工学研究科
40
6.結論
目的より以下のことが結論として導き出せた。
①TGF- β1曝露によるHpMCの形態変化の観察及びmRNA測定からのEMT
誘導の確認
10ng/mlTGF-β
1で誘導されるHpMCは
・敷石状の形態から扇平で細長い形態に変化した。
・転写因子Snail
mRNA発現が増加し、誘導後24hで最大の発現を示
した。
mRNAが時間依存的に減少した。
・細胞間接着タンパク質E-cadherin
形態変化を伴うEMT発生の主な3種類の指標を確認することができた
ので、本モデルをEMT発生モデルとして三次元細胞遊走実験に使用し
た。
またこの条件のときその他のmRNAは
・FNレセプターα5-integrinmRNAはTGF-β
1誘導12時間後から発
現を増し, 48時間後でも維持していたo
mRNAはα5-integrinと同じよう
・またコラーゲン分解酵素MMP-2
に12時間後から発現が見られ、48時間後においても発現が維持さ
れていた。
このことから、
EMTを起こしたHPMCはα5-i山egrinを強発現してお
り、FNをリガンドとして結合する事が可能であることが明らかとなったo
またコラーゲン分解酵素であるMMP-2が問質層中のコラーゲンを分解
することが可能であることが確認できた。
②EMT誘導中皮細胞が化学層化性因子の濃度勾配を認識して遊走すること
をin vitro実験系で実証
・三次元遊走モデルを用い、化学走化性因子としてBSAを用いると、
下層-遊走した細胞数はcontrolと比較して1.2倍ほどであった。
・一方、化学走化性因子としてFNを用いると、下層-遊走した細胞
数はcontrolと比較して2.1倍ほど増加した。
・細胞増殖試験から本実験で使用したEMT発生モデル及び三次元遊
走モデルで細胞数は変化することは無かった。
EMTを起こしたHPMCは、周囲に存在する化学走化性因子を認識して
遊走していることが明らかとなった。
41
③EMT誘導HPMCの腹膜問質層-の遊走の原因を証明
TGトβ
1誘導によりHpMCはα5-integrinmRNAとMMP-2
強発現を起こしている
・
・
mRNAの
BSAの濃度勾配に対して遊走細胞数の有意差は見られなかったもの
の、
FNの濃度勾配においては大きな有意差が見られた。
・またFNの濃度勾配作成三次元遊走モデルにおいて、
るRGDSにより大きな阻害が見られた。
FN阻害剤であ
EMT誘導HPMCの腹膜問質層-の遊走の原因としてFNが関与している
ことが考えられる
④従来の三次元遊走評価法をより改良した新しい三次元細胞遊走装置を誌作
今回作成した新規三次元細胞遊走装置は、化学層化性因子の濃度勾配を
考慮したものである。本実験の結果から、
EMT誘導HPMCは化学層化性因
子特にFNを認識し、そのFNの濃度勾配に従って遊走細胞数が増加すると
いうものになった。
このことから今回作成した新規三次元細胞遊走装置は、より生体に近い
条件を作り出すとともに、以前までのものと比べると、細胞遊走メカニズ
ムの解明をより可能にするものと証明できた。
最後に腹膜透析時におけるHPMCのEMT発生についてまとめると、HPMC
はTGF-β1によりSnail
mRNAを発現し、それによってE-cadherinが減少し
た。さらにα-SMAを発現することにより形態が扇平状に細長く変化した。
α5-integrinを強発現し、問質中に存在するFNを認識する事で、濃度勾配に
沿って濃度の高い方-と遊走した.これによりEMTを起こしたⅠ廿MCは問
質中に存在する化学走化性因子を認識して問質-遊走し、問質線維症を引き
起こす原因の一つとなることが示唆された。
:.車人ノ、;I:人Jt;':院 J'_苧桝JJ/ti約
42
7.今後の展望
・細胞遊走に関与が考えられるシグナル伝達物質の変化の測定
細胞遊走に関与が考えられているシグナル伝達物質、FAKやERRが今
回行った三次元遊走実験条件下で活性化が起きるかどうかウェスタンプ
ロットで測定する。
・阻害剤を利用したシグナル経路の解明
前記に書いたFAKやERRなどを活性化することで細胞の遊走能が増
加することは知られているものの、どのような経路を通ってシグナルが
伝達するかについてはまだ十分にわかっていない。そこでさまざまなシ
グナル阻害を行い、シグナル経路を同定する。
様々な化学走化性因子(礼-8など)を用いて三次元遊走実験
様々な化学走化性因子を用いる事で、EMTを起こしたHPMCに対して
より強く機能する化学走化性因子を検証する。
43
8.参考文献
1坂井建雄,河原克雅著.カラー図解人体の正常構造と機能口腎・泌尿
器.日本医事新報社1999
2
3
GaryA.T,
1998
etal.カラーで学ぶ解剖生理学.医学図書院MYW,
飯田喜俊,二瓶宏,秋津忠男,梼原美治編.
◎EBM血液浄化療法.樵
式会社金芳堂2000
4
平原一郎,他.中皮細胞のepithelial-to-mesenchymal
transition(EMT)による
腹膜肥厚の促進.腹膜透析2007東京医学社
5
Gotloib L,
良. Gokal,
In:
Dordrecht:
6
et al. Functional
Selgas
et
R,
et al.
(eds) Textbook
al
Kluwer
of the peritoneum
structure
2000;
dialyzlng
a
Dialysis
Peritoneal
or
membrane,
Second
Edition.
37-106
Epitbelial-to-mesenchymal
role in the response
as
transition
or the mesotbelial
to dialysis. NephroI
orthe peritoneum
Dial
cell-its
Transplant
2006;
21:ii2-ii7
7
Jimenez-Hefferman
丘broblast
J.A,
9
10
Yanez-Mo
Bucala
et al.
Luiz
Aroeira,
in Peritoneal
Therapeutic
Dokic
D,
et
et
cells. Dev
M,
Zeisberg
epithelial-to-mesenchymal
14
2003;
9: 964-968
Maszi
A,
actin
et al. Central
expression
during
et
al.
∫Am
Soc
role for Rho
that
subpopulation
Transition
Dev
actlVlty
Pathologic
Nephro1
Analysis
2005;
BMP-7
counteracts
reverses
Expression
During
132-142
epithelial-mesenchymal
489-504
chronic
in TGF-betal-induced
epitheliaトmesenchymal
and
and stabilizing lnterCellular adhesion
299:
and
Peritoneal
Significance
234:
stimulated
and
1 8:2004-2013
Gene
Of
Dyn
2007;
Bio1 2006;
transition
403-413
leukocyte
new
Patients:
inhibits TGFbeta
in epICardial mesothelial
13
Dialysis
Rbo
348:
Mesencbymal
transformation.
by modulating
peritoneal
Epithelial-to-Mesenchymal
2003;
a
in
1: 7ト81
al. Microarray
al. VCAM-1
transfbmation
∫Med
to
lnteⅣentions.
Epithelial-Mesenchymal
12
1994;
Med
al. Epithelial
et
Failure
D,
Engl
and
of
247-256
and
Circulating fibrocytes difine
tissue repair. MoI
LaGamba
444:
Dialysis
Cells. N
mediates
Potential
2004;
characterization
tissue
peritoneal
Arch
Peritoneal
al.
orMesothelial
Membrane
ll
et
Transition
S.
normal
fibrosis. VlrChows
M,
R,
Immunohistochemical
al.
in
subpopulations
dialysIS-induced
8
et
TGF-betal-induced
renal
alpha-smooth
transition. Am
rj''J[t z'.ノ、i::研'光村
:_求人'、If・'人ノi::
injury. Nat
∫ PhysioI
Med
muscle
Renal
44
Physio1
15
2003;
TianY.C,
enhance
284:
F91トF924
et
al. Epidermal
HK-2
cell
Res
2007;
313:
Ha
H,
al. High
et
fibronectin
and
through
a
mlgration
factor-betal
transforming
growth
synerglStic
increase
of ERR
and sustained activation
metalloproteinase
16
factor
growth
signaling
of
matrix
Exp
pathway.
Cell
2367-2377
PKC
glucose-induced
by
synthesis
peritoneal
activation
Kidney
cells.
mesothelial
1 and
TGF-beta
mediates
tnt
59:
2001;
463-470
17
0ffner
F.A,
H, et al. Early
0kada
J Physio
19
1997;
L.R,
White
factor-beta synthesis by human
growth
cells. Induction by interleukin-1.
meso血elial
18
Transforming
et al.
during
epitbelium
148:
1679-1688
transfわrmation. Am
F563-F574
al. The
et
∫Patbol, 1996;
in epitbeliaトmesenchymal
role orFspl
273:
Am
peritoneal
characterization of alpha5-integrin
injury. Am
renal
∫ PhysioI
expression
on
Pbysio1
2007;
Renal
tubular
292:
F567-F576
20
Strutz
F, et al. Role
transformation.
21
J.M,
Fan
et
Kidney
tnt 2002;
al.
Interleukinll
transdifferentiation
22
through
mechanism
in vitro. Am
01dfield
M.D,
et
L,
et
al.
a
induces
tubular
transformlng
growth
Dis 2001;
P68
helicase
end
the
via
2001;
cause
products
receptor
108:
for
advanced
1853-1863
PDGF-induced
mediates
Axin
transition by displacing
mesenchymal
factor-betal-dependent
glycation
J Clin Invest
RNA
epithelial-myofibroblast
37: 820-831
transdifferentiation
(RAGE).
in epithelial-mesenchymal
1714-1728
:
Advanced
al.
glycation end products
Yang
61
J Kidney
epithelial-myofibroblast
23
factor-2
of basic fibroblast growth
epithelial
beta-catenin. Cell 2006;
&om
127.・
139-155
24
Liu Y;
al. Epitbelial
et
slgnificance,
Nephro1
to
molecular
2004;
transition in renal丘brogenesis:
mesencbymal
mechanism,
therapeutic
and
intervention.
pathologlC
JAmSoc
15: 1-12
25竹縄忠臣,他.実験医学3細胞極性と細胞運動.羊土社2001
26
GreiliI唱D,
&om
et
collagenous
al. Fibronectin
stroma
provides
a
conduit
into fibrin clot provisional
fbr丘broblast
transmlgration
matrix. J Cell Sci 1997;
1 10:
861-870
27
Bates
R.C,
et
al. TranscrlPt10nal
epitbeliaトmesencbymal
aggressive
transition
colon carcinoma.
activation
de血es
J Clin Invest
a
2005;
of
novel
integrln
beta6
prognostic
during
indicator
the
of
1 15: 339-347
45
28
Strizzi
L,
et
hyperplasias
and
Grinnell
f:
B,
neutrophil
mlgration
3 1 Takahashi
Zhuang
S,
epithelial
33
al. Role
et
an
gland
matrices:
a
interleukin18
of
or
transgenic
lnvest
epidermal
2006;
or
3 12: 86-94
kinases in stimulating
1999;
79:
Res
1403-1413
1995;
factor
growth
Am
mlgration.
and
E貫P Cell Res
Cardiovasc
endothelial monolayer.
Of the
study mlgration
endothelial interleukin-8
of endogenous
al. Requlrement
to
model
phosphorylated
gels. Lab
tbrough丘brin
cell proliferation
characteristic
MMTV-Cripto-1
new
in three dimensions.
et al. Effects
across
mlgration
287:
et
M,
collagen
populations
Schnyder
30
from
a
266-276
201:
Nested
et al.
buman丘broblast
32
2004;
is
transition
mesenchymal
in mammary
tumors
∫Cell Physio1
mice.
29
Epithelial
al.
J PhysioI
on
neutrophil
29: 670-675
in renal
receptor
Renal
Physio1
2004;
F3651F372
Becker
P.M,
VEGF
in pulmonary
2001;
et
al. Differential regulation
of diverse physiologlCal
endotbelial cells. Am
∫ PhysioI
Cell
Lung
to
responses
MoI
Physio1
L1500-L1511
281:
34金森誠司著.ヒト服膜中皮細胞の傷害修復過程におけるヒアルロン酸合
成酵素HAS2
mRNA発現とヒアルロン酸産生の研究.平成17年度三重大
学大学院工学研究科博士前期過程分子素材工学専攻修士論文2006
35
Leung
J.C, et al. Glucose
human
2005;
36
peritoneal
20:
downregulate
ce11s・・ the role of VEGF・
mesothelial
et
al. Cooperation
between
essential f♭r TGF-betaトinduced
Cel1 2006;
37
products
ZO-1
in
NephroI
expression
Dial
Transplant
1336-1349
D,
Medici
degradation
snail and
LEF-1
factors
transcrlPtlOn
transition. MoI
epithelial-mesenchymal
is
BioI
17: 1871-1879
Itoh F, et al・ The
is sufficient
TGF-beta/Smad
FYVE
for
in Smad
domain
localization
of
signaling. Genes
anchor
SARA
in
Cells 2002;
f♭rreceptor
early
7: 321-33
(SARA)
activation
endosomes
regulates
and
1
38坂野真利著.活性酸素種による細胞間結合タンパクの非局在化に関する
研究.平成18年度三重大学大学院工学研究科博士前期過程分子素材工学
専攻修士論文2007
39竹縄忠臣編.細胞骨格と細胞運動その制御メカニズム.シュプリンガ
-
40
Yang
・フェアラーク東京株式会社2002
J, et al. Dissection
transition
and its implications
key
of
events
in tubular
to
epithelial
in renal interstitial丘brosis.Am
myofibroblast
∫Patho1
2001;
159:
1465-1475
41
Yang
L,
et
al. Gene
targetlng
Of Cdc42
and
Cdc42GAP
:,有人′、芦人ノ';::院 卜J、i,I:研JJjt}t村一
affirms
the
critical
46
involvement
in
Cdc42
of
proliferation in primary
filopodia
mouse
induction,
directed
fibroblasts. MoI
embryonic
mlgration,
BioI
Cell
and
2006;
17.I
4675-4685
42
Garcia-Vicuna
R,
CXC
and
matrix
prolifTeration, and
synoviocytes
al. CC
et
cbemokine
receptors
metalloproteinase
&om
rheumatoid
al.
Rho-associated
arthritis
mediate
by
production
Arthritis
patients.
migration,
fibroblast-like
Rheum
2004;
50:
3866-3877
43
Zhao
Z,
et
differentiation and mlgration.
44
R,
Kalluri
Flessner
Renal
46
M.F;
et al. The
288:
2001;
12: 2787-2796
Chen
S, et al. High
112:
in
role
cell
endocardial
275.・ 183-191
its implications
transition and
for
1776-1784
in intraperitoneal
bamier
Am
therapy.
J PhysioI
F433-F442
and
in peritoneal
is mediated
Bio1 2004;
transport
S, et al. Glucose
expression
47
2003;
Physio1 2005;
0gata
Dev
a
play
al. Epithelial-mesenchymal
et
丘brosis. ∫Clin lnvest
45
kinases
mesothelial
cells
glucose-induced,
B
via NF-kappa
alter basic
prednisolone
and
fibroblast growth
fibroblasts. JAmSoc
and
Am
Cell
J PhysioI
Nephro1
fibronectin synthesis
endothelin-dependent
AP-1.
factor
Physio1
2003;
284.・
C263-C272
48
49
IsajiT,
et al.
N-glycosylation
subunit
is essential for
surface,
and itsbiologlCalfunction.
臥,
Inagi
and morphologlCal
Lett
dialysis. FEBS
by
51
J.X,
et al. The
1999;
2006;
103: 623-630
Han
S,
et
52
Gao
M,
Res
et
2003;
Jose
product
factor
of
receptor
:
33258-33267
methylglyoxal
in peritoneal
the
enhances
cells: role in the
in peritoneal
with
COX-2
gamma
in epithelial ovarian
up-regulation
gene
or
Snail.
expression
in human
cancer
peroxisome
cells. Clin
cancer
cervical
Onco1
Gynecol
through
cells
9: 4627-4635
al. Structure
V, et al. A
281
the cell
on
expression
of peritoneal membranes
of invasion
andfunctional
fibronectin type IIIl intermediates. PNAS
53
2006;
alpha5
463.'260-264
is associated
al. Control
proliferator-activated
Cancer
alterations
reinforcement
beta-Estradiol
17
degradation
Chem
of vascular endothelial growth
functional
Ding
J BioI
of the integnn
domain
heterodimerization,
alpha5betal
al. Glucose
et
production
50
of the beta-propeller
synthetic
III4/5) of fibronectin
peptide from
promotes
slgnificance
2003;100:
of mechanically
14784114789
the heparin-binding
stress-fibre and
unfolded
domain
focal-adhesion
Ill
(repeats
formation
in
47
melanoma
54
Jose
V,
Sulfate
Proteoglycans
DeannaLee
2004
57
M,
60
Hirahara
I,
et
Martin
2004;23
ofFibronectin.
Type
III Modules
cell adhesion
and
BioMed
signaling.
Waves
and
2003;33
:30725-3073
Takagi
J et al. Structure
Sambra
D
2008;
doi: 10. 1093/ndt/gfn495
integrlnS
and
in normal
and
tumor
Secretion
Role
for
Induced
A2
by Thromboxane
3-Kinases.
Phosphoinositide
JBC
1
of integrln
in complex
a5bl
fibronectin.
with
EMBO
5
De丘ning
al.
et
ls
Src
orPlatelet
Critical
a
et
B
Jean-Louis
cells in rats.NDT
by
thickening
peritoneal
:7928-7946
Receptor
JBC
2000;
Domain
f♭r
Domain
aMinimal
Adhesion
Synergy
by
Site
149:52ト527
Cation-binding
al. The
Cell
Fibronectin's
&om
f■ibronectin
the
a
Subunit
of Integrln
RecognltlOn
JBC
:24744-24753
Sbiokawa
S. Functional
in EmBryo
Role
Implantation
A
et
al. Crystal
human丘bronectin.
orArg-Gly-Asp(RGD)-Binding
Using
1999;60:
ofReproduction
Sharma
Chondroitin
12:2 1-30
induces
interplay between
L, et al. Two
1 998;273
64
2004;
in tumor
mesothelial
P, et al. The
Zhenyu
a5bl
63
III Domain
Stability orFibronectin
Methylglyoxal
al.
Site-directed Mutagenesis.
62
and
10. 1 186/1477-7827-2-3
;__do主:
2003;22:4607-461
61
lntegrln
a4bl
to the Heparin
Stmcture
Syndecans
et al.
biology. Nature
59
Adhesion
the Mechanical
Variations
mesenchymal-like
58
f♭r Activated
1999;274:135-142
throt唱h Sequence
56
Role
in Cell
C, et al. Tuning
David
2003;371:565-571
al. Cooperative
et
BioCbem
55
Biocbem
cells.
Mouse
and Human
bl lntegrin
on
Decidua.
Biology
1468-1474
stmcture
EMBO
Blastocysts
sites
1999;18:
a
or
heparin-
and integrin-binding
segment
or
1468-1479
:A
illLT(人乍人J?I: L'';i-三!/./、、i,'二研J'Itli
48
9.謝辞
本研究を行うにあたって、研究及び研究生活全般において、適切かつ懇切
なご指導を賜り、公私においてお世話になりました三重大学工学部分子素材
工学科生体材料化学研究室の堀内孝教授、宮本啓一准教授、事務員の村上
節子様に深く感謝いたします。
また本研究を行う上で必要な大網組織を提供していただいた患者様、そし
てそこに関わる諸先生方に感謝致します。
最後に、実験において大きなサポートや、貴重な意見をもらえたMl佐野
晃史君、4年渡部雄一郎君ならびに生体材料化学研究室の皆様に感謝いたし
ます。
49
10. APPENDIX
A-1.細胞培養プロトコル・
・
I
I
・
・
・
・
・
・
・
・
・
・
A-2.細胞の2次元遊走距離の測定(ImageJの使い方)
A-3.AGPC法プロトコル・
・
A-4.RNA定量プロトコル・
・
.
I
A_5.RT_PCRプロトコル・
・
・
I
・
・
A-伝.ウェスタンプロット法・
・
・
・
I
・
・
・
I
・
・
・
・
・
I
・
B-2.PrimerList・
I
・
・
・
・
・
・
B-3.RTnCRprogram・
B-4.統計処理・
・
I
・
・
・
・
I
・
・
・
・
・
・
・
・
・
・
-
・
・
I
I
I
・
I
・
I
・
・
I
・
・
・
I
・
・
・
・
・
I
I
・
・
・
・
・
・
・
・
・
P.$2
・
P.96
P.105
P.107
・
I
I
・
・
・
P.65
P.103
I
・
I
・
・
・
P.63
1
・
-
・
・
.
・
・一・
・
・
・
P.60
・
・
・
-
・
・
・
P.56
I
・
・
・
-
・
・
・
・
I
・
I
・
・
A-A.細胞増殖試験(ImageJの使い方)B-I.滅菌について・
・
・
・
I
I
・
・
・
A-7.三次元細胞遊走モデルの作製・
・
・
・
・
・
P.50
・
・
P.lo島
P.Ilo
A-1.細胞培養プロトコル
A-1-1.培地の調整
<使用機器・器具・試薬>
・
pHメーター(D52-55侶ORIBA)
・吸引ポンプ
10mlシリンジ(TERUMO⑧)
・
・
50mlのシリンジ(TERUMO⑧)
・注射針(TERUMO㊨/NN-1838R)
Millex-GV
(MILLIPORE/Cat.No.SLGVO25LS/孔径o.22
・
I
・
・
・
・
・
strivex GS
(MILLIPORE/Cat. No.SVGSBIOIO/孔径o.22
sTERICUPTMフィルター(MILLIpORE
pm)
pm)
/ SCGVUO5RE)
30mlチューブ(アシスト/543.001S)
o.5mlアシストチューブ(アシスト/72.699S)
Medium
penicillin
199(SIGMAノM2154)
(SIGMAn-0781)
streptomycin
主▲A
▲.車人'、;三:人ノ';三:院
J、jl':研JJ/t'.キ斗
50
Insulin
Transferin
Selenate
(SIGMA/
しGlutamine
1N
FBS
aq
G-7513)
(Wako /196-05635)
aq
HCl
Ⅰ-1844)
(SIGMA/ H-0888)
Hydrocortisone
1NNaOH
(SIGMA/
(Wako /080-08065)
(SIGMA/
F9423)
<各溶液の調製>
O
(牛胎児血清)の不活性化、分注、保存
Fetalbovinesemm
(丑 凍結の状態で送られてきたFBSは37℃で溶かしたo
溶かし終わっ
たら数回振り、55-56℃で、時折振り混ぜながら30分間温めた(不
活性化)0
② 50mlのシリンジで吸い取り、
StrivexGSで、もしくは、
Stericupフ
30
ィルターユニットにてフィルター滅菌し、
mlチューブに20
ずつ分注した。作業はクリーンベンチ内、滅菌操作で行った。
ml
③ 冷凍庫(-20℃)で保存した。
○培地の調整
I
ITS
(InsulinTransferin Selenate)
(液体)を1.5mlアシストチューブに500〃1ずつ分注し、冷
ITS
蔵庫にて4℃で保存した。
(L-Glutamine)
・しG
15ml遠沈管に5mlずつ分注し、冷蔵庫にて4℃で保存した。
・
(Penicillin-Streptomycin Solution)
P-S
5mlアシストチューブに5mlずつ分注し、冷凍庫にて-20℃で保
存した。
・
(Hydrocortisone)
HC
HC
4
mlの無水ethanolで溶解し、 DEPC処理水2
mgを2
mlを加
えて漉過した。 o.5mlアシストチューブに200plずつ分注し、
R.T
で保存した。
① L-Gとp-s各5ml、
ITSとHC各50plを添加量とした。
上記のサプリメントを、
②
10mlシリンジを用いてMillex-GVを通し
てフィルター滅菌し、培地(Medium-199)に添加した。
※pHを調整する場合
③
培地のpHをpHメーター、
1NNAOHaq、
1NHCLaqを用いて調整
51
した。また、
pHは7.2に調整した。
※体内のpHは7.4である。(フィルターにかけるとpHがo.2ほど上昇
するため)
A-1-2.初代培養
<使用機器・器具・試薬>
・
co2インキュベーター(池本理科工業/10-0212)
・
FORCED
I
KUBOTA2010
CONVECTION
(ADVANTEC/DRS620DA)
OVEN
(株式会社久保田製作所収D3132)
・撹拝遠心機(LABINCO/2790001)
・ウオーターバス
・
Vortex
・滅菌済手術器具一式
25cm2培養フラスコ(住友ベークライト/MS-21050)
・
・
・
15ml遠心管(住友ベークライト/MS-56150)
50ml遠心管(住友ベークライト/MS-56500)
・遠心管立て
10mlピペット(住友ベークライト/MS-66100)
・
・ピペッタ・タイマー
I
199
Medium
(SIGMAM154)
(SIGMAD8537)
・
PBS
・
TRYPSIN-EDTA
・
FBS
SOLUTION
(lox) (SIGMA/T4174)
(SIGMA/F9423)
<組織・細胞の入手>
本研究で使用したHpMCは、予め本学医学部倫理委員会にて患者組織の
採取につき承認を得、かつinfbrmed
consentを得た患者より頂いた大網組織
に由来する。
Ⅰ
組織の切断
①オートクレーブ滅菌処理した金属トレイに、滅菌済み(新品)のPBS
を組織が浸る程度の量だけ入れる。
②すぐさま組織を浸し、切断箇所の上下を滅菌糸で固く縛り、縛った箇
「 '、;::桝''JTf
:.辛)こ一';I:人二J'[[二F,(LT
i:i,
52
所の間を滅菌済みハサミにて切断する。
③同様の操作を繰り返し、
3つ前後の親指大の切片に切り分ける。
基本操作
Ⅲ
①
25
c招培養フラスコに10%FBS/M-199を5ml加えてプレインキュベ-
卜しておく。
② 50ml遠心管に、
5.0%トリプシン/2.0%EDTA溶液2mlとpBS18mlを加
え、 o.5%トリプシン/2.0%EDTA溶i夜仲BS
(以下トリプシン溶液)を
作製する。
③用意した50ml遠心管にトリプシン溶液20mlを加え、親指大の大きさ
にした大網組織を入れて、
37℃/20分間摸拝遠心機により摸拝しなが
らインキュべ-トする。
④授拝した大網を遠心管から取り出し、次の遠心管に入れ、
行う。また、大網を取り出した遠心管は50G
5分間遠心分離を行う。
⑤上澄を抜き取りcellpelletを作る.
⑥再び上澄を抜き取り、
(遠心半径10cm750叩m)
50%FBS刀M-199を2ml加え、
(遠心半径10cm750叩m)
をかけて、再び50G
③の操作を
vortex
5分間遠心分離を行う。
10%FBS/M-199を1ml加えて10回のピペッテ
ィングにより細胞懸濁液を作成する.この懸濁液を(Dで調整した25
cI丘培養フラスコに播種し, 37℃の5%CO2インキュべ-ト内にて培養
する。
⑦ ③-⑥の操作を4回線り返す。
A-1-3.継代培養
<使用機器・器具・試薬>
I
・
co2インキュベーター(池本理科工業/10-0212)
KUBOTA2010
(株式会社久保田製作所収D3132)
・ウオーターバス
・
Vortex
・吸水ポンプ(日機装侶R-21B)
75cm2培養フラスコ(住友ベークライト/MS-21250)
・
・
・
・
25cm2培養フラスコ(住友ベークライト/MS-21050)
6穴プレート(住友ベークライト刀MS-80060)
¢35mmシャーレ(住友ベークライトパMS-80240)
53
15ml遠心管(住友ベークライト/MS-56150)
50ml遠心管(住友ベークライト/MS-56500)
遠心管立て
10mlピペット(住友ベークライト/MS-66100)
ピペッタパスツールピペット
タイマー
チップ各種
セルスクレーパー(住友ベークライトrMS-93170)
Counting
(Erma/03-202-1)
chamber
血球計算版
199
Medium
PBS
(SIGMA仙蛇154)
(SIGMAM8537)
TRYPSIN-EDTA
FBS
SOLUTION
(lox) (SIGMA/T4174)
(SIGMA/F9423)
細胞の洗浄
①
培養フラスコの蓋を開け、口元を加熱殺菌する。
②
滅菌済みパスツールを加熱滅菌し培養フラスコ内に挿入し、培養液
を吸引する。
③
pBS溶液5ml/25cdフラスコ又は10ml/75cI丘フラスコを添加し、前後
左右に振り洗浄する。
④
滅菌済みパスツールを加熱滅菌し培養フラスコ内に挿入し、洗浄液
を吸引する。
Ⅱ
培養細胞の剥離方法(培地:mesoの場合M-199
、丘broの場合
F-12)
①冷凍保存してあるトリプシン1mlにPBS9mlを加え、10倍希釈する。
(以下トリプシン溶液)
②トリプシン溶液5ml/25cI丘フラスコ又は10ml/75c遥フラスコを加えるo
③顕微鏡で細胞が浮いてきたことを確認後、フラスコを軽くたたく。
④ FBSlml/25
cI丘フラスコ又は2ml/75cdフラスコを加える。
⑤滅菌済みセルスクレーパーを培養フラスコ内に挿入し、細胞を剥離す
「.
[L.
,T・:)( I?I:人一?'二!;`;tI?:桝''Jt村
54
る。
⑥滅菌済み10mlのピペットで培養フラスコ内の細胞懸濁液を吸引し,
15ml遠心管に入れる。
⑦ 50G (750rpm5分間)遠心分離する。
⑧培養フラスコに10%FBS/培地を入れ,
ト内にて放置する。
37℃の5%CO2インキュべ-
(75cm2フラスコ-15m1
6穴プレート-2ml/
穴)
⑨ ⑥で遠心分離した上澄みをピペットで吸引する。
⑩ 10%FBS/培地を1ml又は3mlを遠心管に沈殿している細胞に加え、
細胞懸濁液を作成し、
10回程度ピペッティングを行う。
⑫細胞懸濁液を取り、
⑦で用意しておいた培養フラスコに播種し、 37℃
のCO2インキエペ-ト内にて培養する。
プレート-200〃
(75cm2フラスコ-1m1
6穴
1/穴)
55
A-2.細胞の2次元遊走距離の測定(ImageJ)
【下処理】
① 「ImageJ」を選択し、起動させる。
② 「File」で目的の写真を選択する。
EdT
h3g8
FhcesS
ATd†yZ8
PALgれI
WrLd⊂■
Jt[p
8かl卜
-hLへl耶:′一言■nrS・LJLLW,P
こで■ヽ SJPE!PI
OpeTI
RecerL
C¶◆W
⊂Lr7◆S
S8咋
Sa>d
AB
Reveyt
CW◆只
Setup
Pi哩e
帆.
Ctn◆P
呈Lril
③ rlmage」を選択し、
「Adju8t」
-
「Threshold」を選択する。
I/u;;:八二■l r;';t
r:r!:
1
1
1=
′jt手
56
㊨ 「Threshold」でピクセル数の上限と下限を設定する
*
目的の細胞が赤の点で固まれるように設定すること
*
またなるべく細胞の教を少なくすること
以上が前処理になる。
*
先ほどの「Thresbold」で「Apply」でも実行できる
57
【実行】
① 下処理が終わった画像を、
②
「Analyze」
-
「SetmeasurementJを実行するo
「Setmeasurement」において「Center&MassJをチェックするD
)(I,: 】こンl;1
r..二)/;;=
58
,二糾1L:手:r-
⑤ 「Analy21e」を選択し、
*
「Analyzeparticle」を選択する
Sizeを1000・10000に設定する
⑥ ⑤を実行後、目的の細胞のⅩ、
Y値を記録する
59
A-3AGPC法プロトコル(6穴プレートの場合)
<原理>
RNAを構成するリボ-スは、
DNAのデオキシリボ-スに比べて2位
の炭素に水酸基が1つ多くある。このためDNAに比べて、次のような条
件下では親水性が増すことが期待される。
核酸は、中性条件下ではリン酸基が解離して負の電荷を帯び、この部
分が水和することによって高分子コロイドとして水中に分散している0
ところが酸性条件下では、リン酸基の電離平衡が会合の方向に傾いてこ
の部分のチャージが失われるために、親水性が低下する。この状態でフ
ェノール処理を行うとDNAは疎水性に勝るフェノール層に分配される
が、 RNAはリボ-スの水酸基があるため水屑に分配される。
AGPC法と
はこの性質を利用して、RNAを抽出する方法である。
(GTC)でRNaseを失活させ
thyocyanate
一般的には、まずGuanidium
ると同時に細胞のタンパク質を可溶化する。そして液性を酸性に保ちな
がらフェノール処理を行うと、タンパク質や脂質がフェノール層や中間
層に移行するのみならず、
DNAもフェノール層に分配される。こうして
得られた水屑(GTC層)にはRNAのみが溶け込んでいるので、これを
アルコールで塩析することでほぼ純粋なRNAが得られるわけである。本
実験では,市販のRNA-BeeTM
ISOLÅrION
OF
RNAと呼ばれる試薬を用
いてRNAの抽出を行っているが、その原理は基本的に同じである。
<RNA
・
DNAを扱う上での注意点>
細胞は原則的に受精卵が持つ全ての種類の遺伝子を持っているが、細
胞の種類・状態に応じて、特定の遺伝子(DNA)を選択的に活性化して
機能を発現している。この仲立ちをするのがmRNAである。したがって、
細胞の状態をフレキシブルに変化させるためには、いつまでも古い
mRNAが残っていては具合が悪い。そこで細胞や組織液中にはRNAを速
やかに分解する酵素(Nase)が豊富に含まれている.我々がRNAの分
析を行う際には、このRNaseによるRNAの変性・分解が最大の障害と
なる。
RNaseは抽出する細胞の他、我々の汗・唾液・皮膚など様々な所
に存在し、また熱に対する安定性も極めて高く、なかなか失活しない
三重人学大学院
工学研究科
60
(121℃の高圧蒸気滅菌にかけても活性が残る)。従って実験の成否はこ
のRNaseの作用をいかに防ぐかにかかっている。このことを踏まえ、今
後RNA、
DNAを扱う全ての実験において以下の点に気を付けること。
○各チューブ・チップは、素手で触れたものは使用しないこと。
○滅菌のために各チューブ・チップを詰め込む際は、ゴム手袋を着用
し素手では絶対に触れないこと。
○使用するガラス器具は乾熱滅菌(RNase丘ee)を行うこと。
○実験中は必ずゴム手袋(片手で良い)、マスク、帽子、白衣を着用
し、無駄な会話は避けること。
○
ゴム手袋を着用した側の手は、サンプルを扱うこと以外には使用し
ないこと。
○サンプルに直接触れるものを素手で扱わないこと。
○サンプルを吸う際は、常に細心の注意を払うこと。
また、
AGPC法に特有の注意点として、以下の点に気を付けること。
○高速遠心機は予冷運転を行っておくこと
○操作は全体的に素早く行うこと
<使用機器・器具.試薬>
・日立微量高速遠心機(日立/cR15B型)
DNA
EQUIPMENTID/
mini (IIETO LAB
・
・
・
Mm
872146)
(COSTE…VSS-06618)
CENTRIFUGE
Vortex
ウオーターバス
・
1.5mlチューブ(アシスト/72.690S)
I
・チューブ立て
・マイクロピペッタ-各種
・チップ各種
・
・
タイマー
crashice用発泡スチロール
pMILM⑧
・
(Pechiney Plastic Packaging,
Inc.仲M-996)
・注射針(TERUMO⑧/NN-2432R)
・
RNAIBeeTM
ISOLATION
OF
RNA
(Tel-Test,Inc./CS-104B)
61
cblorofbrm-isoamylalcoho1
isopropanol
24: 1
(SIGMAノC-0549)
(SIGMA/Ⅰ-95 16)
(SIGMAD-8537)
PBS
75%EtOH/DEPC処理水溶液(無水EtOHをDEPC処理水で希釈)
(Wako/321-00025)
-無水EtOH
DEPC処理水
→
Crash
Diethylpyrocarbonate
1 1)
(CALBIOCHEM⑧/2987
ice
<AGPC法(前半)
>
①細胞に刺激を与え、
6穴プレートから培地を吸引し、
PBSを1ml/well
入れて細胞を洗浄する。
※後の行程を考慮し、
①
3穴または4穴毎に行うのが良い。
洗浄後pBSを吸引し、素早くRNA-BeeTMを1ml/well入れる。
②
2連にしたピペットチップの先端でwell内の細胞を剥がすように掻
いた後、
10回程度ピペッティングを行い、その懸濁液を1.5mlチュ
ーブに移し入れる。
※ ②と③の操作を5分間で行うこと。
※チップはwell毎に交換すること。
③
チューブに入った各サンプルにchlorofbrm
-
isoamylalcobo1
24:1を
200〃1ずつ加え、しっかりと蓋を閉めた後15回程度上下に振り混
ぜる。
④
室温(良.T)にて15分間静置する。
⑤
12000G/4℃/15分にて遠心分離を行う。
⑥
3層に分離するので、最上層の溶液(透明な水屑、ここにRNAが含
まれる)を液面近傍よりそれぞれ350〃1ずつ採取し、別に用意した
1.5mlチューブに移し入れる。
⑦ 移し入れた各サンプルにisopropanolを500〃lずつ加え、
vortexに
て撹拝した後すぐに-80℃の冷凍庫に移し一晩保存する(OverNigbt)0
:.車人′?I:人Inf!i';t
「ノ、;・[:桝先手:i
62
<AGPC法(後半)
>
【全て氷上操作】
①サンプルを解凍し、
12000G/4℃/10分にて遠心分離を行う。
※ このときチューブの継ぎ手を外側に向けて遠心を行うこと。
②チューブを傾け、ある程度の上澄を慎重に取り除く。
※沈殿は肉眼では確認出来ないので、チューブの継ぎ手側に沈殿があ
ると想定して上澄を取り除くこと。
③各サンプルに75%EtOHのEPC処理水溶液を500I1
1ずつ加えvortex
にて撹拝した後、
10000G/4℃/5分にて遠心分離を行う。
④チューブを傾け、より慎重に上澄をぎりぎりまで取り除く。
⑤チューブの口に2.5cm四方程度のPMILM⑧を張り、注射針にて
10個程度の穴を開ける。
※ pMILM⑧は絶対に素手で触らないこと.
⑥ DNAminiにサンプルをセットし、減圧乾煉を行う。
※サンプル残量によって乾燥の度合いが異なるため、乾燥し過ぎない
ように様子を見ながら行うこと(目安:1セット3-5分)。
⑦各サンプルにDEPC処理水を10FLlずつ加え、
MINI
vortexによる撹拝と
CENTRⅣUGEによる軽い遠心を2回線り返す.
※減圧乾燥の際にRNAが陰圧で壁に付着する場合があるため、念入り
に行うこと。
⑧ウオーターバスにて55℃/5分加温後、冷蔵庫にて4℃に冷却する。
⑨得られたサンプルはRNAの定量後、
RT-PCR法に使用する.
A-4RNAの定量プロトコル
<意味>
ここでは、分光光度計を用いて吸光度oD260およびOD28.の測定を行い、
その値を基に全サンプルのRNA濃度を一律に揃える。これにより、この
後に行うpcRの反応生成物に定量性を持たせることが出来る。というの
も、 RT-PCR法はmRNAを鋳型にしてDNAの合成・増幅を行い、得られ
たDNAの定量を行うため、最初の鋳型mRNAの量にばらつきがあると
最終的なpcR反応生成物畳もばらついてしまい、定量性を失ってしまう
からである。
63
<注意点>
核酸(この場合はRNA)由来の吸光度oD26。とそれ以外の物質(タン
パク質など)由来の吸光度oD28.の比(OD260/OD28.)は、サンプルのRNA
純度を表しており、一般的に1.5以上が望ましいとされる。但し、いくら
比が大きいとしても吸光度が低いとRNA濃度は低くなってしまうため、
理想的なサンプル濃度o.2-0.5〃g/FLlにするためには、少なくとも
oD260-0.085以上は必要である。また、逆にサンプル濃度が高過ぎる場合
は希釈して用いること。
<使用機器・器具・試薬>
I
smart
I
cuvette
・
MINI
・
Vortex
I
SpecTM
3000
Spectropbotometer
(BIO 久AD/170-2501)
(BIO 久AD/170-2505)
(COSTER/MVSS-06618)
CENTRⅣUGE
o.5mlチューブ(アシスト/72.699S)
・チューブ立て
・マイクロピペッタ-各種
チップ各種
t
(株式会社クレシア/62011)
・キムワイプ⑧s-200
・洗瓶(脱イオン水)
o.1NHClaq
(Wako/080-08065)
I
I
DEPC処理水(CALBIOCHEM/298711)
<RNAの定量>
① AGPC法により得られた各サンプル10〃1より1〃1ずつ採取し、サ
ンプル数分用意したo.5mlチューブの底部にそれぞれ入れる。
② DEPC処理水を59FLlずつ①のチューブに加え、
vortexにて撹拝後
MINI
CENTRⅣUGEにて軽く遠心を行う。
③前日よりHCl
aq (0.1N)に浸け置いたCuvetteを、脱イオン水にて
よく濯ぎ水気を切る。
④マイクロピペッタ-を用いてCuvetteにDEPC処理水を60[Ll入れ、
Smart
SpecTM
3000
Spectrophotometerにセットし、零点補正(Read
:.
卜Jli:I:研JJ]t村
FT7:八一';:二人'';-I:院
64
Blank)を行う。
⑤マイクロピペッタ-を用いてDEPC処理水を抜き取り、
定サンプルを60FLl入れSmart
SpecTM
3000にセットし、
1つ目の測
3回吸光度
の測定(ReadSample)を行うo
(歩測定し終えたサンプルを、マイクロピペットを用いて抜き取り元の
チューブに戻す(測定のやり直しが出来るようにするため)0
⑦ cuvetteを脱イオン水にてよく濯ぎ水気を切る。
⑧ (9-⑦をサンプル数分繰り返す.
⑨測定結果が異常であったサンプルを再測定する。
⑩得られたデータを出力し、データの処理を行う。
<データの処理>
得られたデータより、次の2つの計算を行う。
i)各サンプルの濃度
並)全サンプルの濃度を最低のサンプル濃度に合わせるために必要
なサンプルとDEpC処理水の量
それぞれに必要な計算式を以下に示す。
サンプル濃度(pg/p
1
)-(OD260)×(換算係数)×(希釈倍率)
RNAの最低量(pg)-最低のサンプル濃度(pg/pl)×サンプル残量(pl)
必要サンプル量(pl)-RNAの最低量(pg)/希釈対象のサンプル濃度(pg/pl)
必要DEPC処理水量(pl)-サンプル残量(pl)一必要サンプル量(pl)
※換算係数はRNA測定の場合o.o4
(jig/FLl).
※本実験における希釈倍率は60倍。
※本実験におけるサンプル残量は、
AGPC法にて得られたサンプル10
〃1からRNAの定量に用いた1〃1を差し引いた9〃1。
A-5T-PCRプロトコル
<原理>
pcRは微量なDNAの存在を非常に鋭敏に検出できる方法であり、こ
の方法を応用すれば極微量のmRNAの発現を検出することができる。し
65
かしPCR法に用いるDNA合成酵素は、
mRNAを鋳型としてDNAを合
成する活性を持たない。その間題の解決方法として重要となる反応が逆
転写反応である。つまり、逆転写酵素を用いてmRNAを鋳型としたCDNA
の合成反応(逆転写反応)を予め行えば、そのcDNAを改めて鋳型とし
てPCRを行うことができる.これがmRNAの発現をPCR法によって検
出する方法、すなわちRT-PCR法である(Fig.A-5-1).
Fig.A-4-1
A-5-1.
Reverse
RT-PCR法
Transcription
<原理>
細胞から発現したmRNAにPrimer
(RandomHexamer)を結合させ、逆
転写酵素を用いてmRNAに相補的な1本鎖DNA
:.車人J'i,I:大`芋:院
l
/、デ:研JJ)tji村
(CDNA)を逆転写反応
66
により合成する。この時点ではDNAとRNAが繋がった2本鎖の状態で
あり、安定性が高いため、
pCRの熱変性(denature)時に十分に1本鎖に
ならないことがある。そのため、DNA鎖と対を成しているmRNA鎖を、
RNA分解酵素(RNase)を用いて分解する。これにより完全長cDNAを
合成することができる(Fig.A-5-1)0
<注意点>
注意すべきことは、とにかくRNaseの混入を防ぐことである。詳しく
はA13AGPC法プロトコルを参照のこと。また、
RT法に特有の注意点と
して以下の点に気を付けること。
O
TEMP
PROGRAM
CONTROL
SYSTEMにRT
program
01/02を予め入
力しておくこと。
O
Reaction
Tube
with
Capは必ず専用の滅菌缶に入れ滅菌しておくこ
と。その際も素手では触れないこと。
O
ReactionTubewitbCapにラベルする際は、分かりやすく、かつ消え
ない箇所に記入すること。
O
ReactionTubewitbCapは他のものと比べて脆く、力を入れ過ぎると
破損する恐れがあるので注意すること。
<使用機器・器具・試薬>
・
・
・
I
TEMP
pROGRjM
MINI
CONTROL
SYSTEM
(ASTEC仲C-708)
(COSTER/MVSS-06618)
CENTRIFUGE
Vortex
Reaction
Tube
with Cap
(MicroAmp伽801-0540)
・チューブ立て
・マイクロピペッタ-各種
・チップ各種
crasbice用発泡スチロール
・
I
・
・
・
・
・
・
(InvitrogeIV48190-011)
Random
primers
dArP
dTTP
(Amersham Biosciences/27-2050-01)
(Amersham Biosciences/272080)
dCTP
(Amersham Biosciences/272060)
dGTP
(Amersbam Biosciences/272070)
buffer (PERENELMERJE12387)
lox
PCR
Recombinant
RNasin⑧
Ribonuclease
lnhibitor
(Promegam25
1 1)
67
Super
ScriptTM
RNase
HI Reverse
DTT
(Invitrogen/YOO147)
Transcriptase
(Invitrogen/18053-01
7)
DEPC処理水(CALBIOCHEM/298711)
Crash
<RT
Master
ice
Mix>
【全て氷上操作】
Table.A-5-1に従い、各試薬を混合しRTMasterMixを作製する。極微量
を扱うため、サンプル数丁度の量ではRTMasterMixが不足する恐れがあ
る。そのためサンプル数が多い場合は1、
と。また、
2サンプル分余分に作製するこ
inhibitorとSuper
Table.A-5-2に後で加えるRNase
ScriptTMの必
要量も示す。
Table.A-5-1
RT
Master
(単位:pl)
Mix
12
18
(+2)
(+3)
EE
14
21
28
7
EE
14
21
28
1
7
ll
14
21
28
dCTP(2.5mM)
1
7
EE
14
21
28
dGTP(2.5mM)
1
7
EEl
14
21
28
10xPCRbuffer
2
14
22
28
42
56
DTT
2
14
22
28
42
56
合計
9
63
99
126
189
252
1
Sample数
6(+1)
9(+2)
1
7
dArP(2.5mM)
1
dTTP(2.5血M)
24
(+4)
RandomPrimers
(100mM)
Table・A-5-2RNase
inhibitorとsuper
1
Sample数
6(+1)
scriptTMの必要量(単位:pl)
9(+2)
12
18
(+2)
(+3)
24
(+4)
RNaseinbibitor
1
7
EE
14
21
28
SuperScriptTM
1
7
EE
14
21
28
2
14
22
28
42
56
合計
:.
)(;・':院
I巨人ノ;::
卜't;I:桝ノ光村
68
< Reverse
TranscrlptlOn
>
【全て氷上操作】
①各サンプルを扱う前にRTMasterMixを作製する。
② RNAの定量結果に基づき、
AGPC法により得られた各サンプル原液
をReactionTubewitbCap内で希釈し、
③希釈後の各サンプル9〃1に、
9〃1とする。
vb什ex
RTMasterMixを9〃1ずつ加え、
による撹拝とMNCENTRIFUGEによる軽い遠心を行う。
④
PROGRAM
TEMP
CONTROL
RT
SYSTEMにサンプルをセットし、
olを実行する(B-5.
RT/PCRprogram参照)0
⑤各サンプルにRNase
inhibitorとSuper ScriptTMを1 〟 1ずつ加える。
program
※予め2液を混合したものを2〃1ずっ加えても良いが、その場合は混
合液の量を4サンプル分程度多めに作製しておくこと。
⑥
PROGRAM
program
TEMP
CONTROL
o2を実行する(B-5.
RT
SYSTEMにサンプルをセットし、
RT/PCRprogram参照)0
⑦作成したRTproductは-20℃の冷凍庫にて保存する(数ヶ月保存可)0
A-5-2_
Polymerase
ChaiII
Reaction
<原理>
pcRの原理は3段階から成るDNAの合成反応を繰り返して行うこと
にある。まず、鋳型となる2本鎖DNAを加熱して変性し、
1本鎖にする
(denature)。次に、増幅したい特定部位のDNA鎖の両端に相補的な2種
類のオリゴヌクレオチドプライマーを反応系に過剰に加えた状態で温度
を下げると、プライマーがDNA鎖の相補的な部位と2本鎖を形成する
(annealing)。この状態でDNA合成基質のデオキシヌクレオシド3リン
酸とDNAポリメラーゼを作用させると、ポリメラーゼはプライマー部位
からDNA相補鎖を合成していく(extension)。ここで重要なのは、最初
の2サイクルでは長さが不定な2本鎖DNAが合成されるが、
3サイクル
目からはプライマー対に挟まれた部位の長さの揃った2本鎖DNAが合成
され、その準が急速に増えていくということである。また、
1回目の合成
反応で生成したDNAは次の反応の鋳型となるため、その名称の通り連鎖
反応的にDNAが合成されることから、20-30サイクルの反応の後には、
もとの鋳型DNA中の必要な部分配列をもつDNA分子を莫大な数得るこ
とが出来る(Fig.A-5-2)0
69
CDNA
Primer
TTTTTT
5
Primer
(antisense)
3'
'塾嘩
AAAAAA
(antisense)
TTTTTT
5'
-叫■■・
Primer
TTTTTT
(sense)
伸長反応
(extensio a)
3'
3
AAAAAA
TTTTTT5,
5
加熱変性
(denatu
)
re
■
■
3'
■
AAAAAA
●
■
n
cycles
■
I
1本鎖へ分裂
理論上2n倍に増幅
TTTTTT
3'
Fig.A-5-2
PCRの原理
<pcRのカイネテイクスとサイクル数>
ここでは、PCRによって得られる反応生成物の量とpCRのサイクル数
の関係について論ずる。
PCRは1サイクルの反応によって目的とする
DNA断片が最大2倍に増えるため、初期鋳型量をⅠ。とすると、
nサイク
ル後の反応生成物量Ⅰは、理想的には次式(Eq.A-5-1)に従って指数関数
的に増加する。
Ⅰ-IoX2n
(Eq.A-5-1)
---
しかしサイクル毎の増幅は、プライマーのアニーリング効率やDNA鋭
の伸長反応の効率が100%ではないことなどの要因によって、必ずしも2
倍にならない。このことを考慮すると、
nサイクル毎の瞬間増加率をE'
とした場合は、反応速度をdI/dnとした次の微分方程式(Eq.A-5-2)で与
えられる。
dI/dn
-
E'×Ⅰ
(Eq.A-5-2)
----・
仮にE'がほぼ一定の値をとると考えて、
E'を定数としてEq.A-5-2を解
くと、単純に次式(Eq.A-5-3)が得られる。
Ⅰ
-
Ⅰ。×En
これにより、
(※E-eE')
・---
(Eq.A-5-3)
PCR反応生成物は増幅率をEとして指数関数的に増加す
ることが期待される。ここで、実際のPCRでも反応生成物がある程度蓄
積するまでは、反応生成物は概ね指数関数的に増加しており、その指数
関数的に増加している部分のデータを元に片対数で直線近似を行うと、
卜乍研'J)t:村
--)T7.:人ノ'i::人一、y二院
70
Fig.A-5-3のようなグラフが得られる。これによると反応生成物が蓄積す
るまでは、
Eq.A-5-3において増幅率Eを一定とみなしたモデルが妥当で
あることが分かる。
Fig.A-5-3
PCR反応生成物量とサイクル数
しかし、反応が進んで生成物量が増加するにつれて増幅率は低下し、
最終的には反応生成物量の増加は頭打ちとなり、一定の値をとるように
なる。反応が停止する要因としては次の5つが挙げられる。
i
)
TaqDNApolymeraseの失活
五)反応基質(デオキシヌクレオシド三リン酸)の枯渇
近)プライマーの枯渇
iv)反応副産物であるピロリン酸による合成反応阻害
Ⅴ)生成したDNA鎖同士の再会合によるプライミングの阻害
i
)に関しては、同じサイクル数でも反応生成物量の増加が頭打ちに
なる場合とそうでない場合があることや、一度頭打ちになった反応液に
さらに酵素を追加しても通常は反応が進行しないことから、通常のPCR
サイクル数(25-30サイクル)においては主たる限定要因にはなってい
ないものと考えられるo
また、
ii) -v)の限定要因については、その
効果が反応生成物量に概ね比例するものと考えることが出来る。
以上のことを考慮すれば、一般的にPCRのサイクル数は30サイクル
前後が良いと言える。もし30サイクルで十分な増幅が見られない場合は、
35サイクル程度までサイクル数を増やしてみると良い。
71
<反応温度と反応時間>
非常に繊細な反応であるPCRは、わずかな温度やサイクル数の変化が
反応の成否を分けると言っても過言ではない。ここでは、
PCRの各段階
における一般的な反応温度と反応時間について論ずる。
i
)熱変性(denature)の温度と時間
熱変性は94℃×30秒-1分を標準とするが、液量が少ないために
反応液の温度がすぐに高温に達するような場合は、
30秒以下でも十
分な場合がある。ちなみに93℃でも十分な場合が多いが、
94℃まで
上げておいた方が無難である。また、反応時間が長すぎると、酵秦
が失活してしまう点にも注意が必要である。
立)アニーリング(annealing)の温度と時間
アニーリング温度はプライマーのTm
(次項
<primer>
参照)に
留意して設定する。時間は通常1分で良い。目的とする標的配列以
外の非特異的配列が増えるようなら、
全く何も増幅されないようなら、
2℃間隔で温度を上げてみる。
2℃間隔で温度を下げてみると良い。
iii)伸長反応(extension)の温度と時間
伸長反応は通常72℃で行う。伸長時間は標的配列の長さによって
異なり、
1000bp以下の場合は1分で十分だが、それよりも長い配列
の場合には1000bpにつき1分の割合で長くする。
< Primer
>
ここにプライマーの選択のための条件を挙げる。自らプライマーを設
計する場合はこれらの点に気を付けなくてはならない。
i)プライマーの長さ
標準的なプライマーは18-28塩基対であり、中でも20-24塩基
対が望ましいとされている。通常この位の長さであれば、鋳型DNA
との特異的なアニーリングに十分であると考えられている。
正) CG含有量
プライマー内でG/C塩基と〟c塩基がほぼ均一になるように設計
することが望ましく、標準的なcG含有量は50%である。また、プ
ライマーの3'端でのÅr含有量が多くならないようにする。
ih) Tm値
Tm
(MeltingTemperature)とは、
PCRにおいてDNAの2本鎖が熱
変性(denature)して1本鎖になる温度のことであり、標準的には55
:.車人J、i三:人′、if:院 1∼.1羊桝`J/LIE村
72
-80℃である。合成する2つのプライマーのTm値は同じであるこ
とが望ましいと考えられている。
Tm値はAとTでは2℃、
CとG
では4℃を目安に計算する。いくつかのプライマーの候補がある場合、
通常Tmの高いものを選ぶ。
iv)プライマー間の相補性
2つのプライマーの、特に3'端の間で相補性のないようにする。
プライマー同士ダイマ(2量体)を形成すると、
pCRの効率が低
下するためである。
Ⅴ)プライマー内の二次構造
プライマー内で二次構造をとらないようにする。これはPCRを阻
害するばかりでなく、プライマーを独自に合成する際の効率にも影
響すると考えられるからである。
<注意点>
注意すべきことは、とにかくRNaseの混入を防ぐことである。詳しく
はA-4.
AGPC法プロトコルを参照のこと。また、
点として以下の点に気を付けること。
O
PROGRAM
TEMP
CONTROL
pCR法に特有の注意
SYSTEMに各プライマーに対応した
PCRprogramを予め入力しておくこと。
○サイクル数を上下させる場合は4サイクル刻みで増減させ、
2サイ
クル刻みで微調整すること。
O
RT
product内に白い沈殿がある場合は、
cDNAが壊れてしまってい
る可能性が高い。そのサンプルでPCRを行っても、結果が得られ
ない場合が多いので注意すること。
O
RT法と同様、
ReactionTubewithCapの扱いに気を付けること。
<使用機器・器具・試薬>
・
・
・
・
TEMP
PROGRAM
MIM
CONTROL
CENTRIFUGE
SYSTEM
(ASTEC/PC-708)
(COSTER爪IVSS-06618)
Vortex
Reaction
Tube with Cap
(MicroAmp/N80110540)
・チューブ立て
t
マイクロピペッタ-各種
・チップ各種
Crashice用発泡スチロール
・
73
dÅrp
(Amersham
Biosciences/27-2050-01
dTTP
(Amersham
Biosciences/272080)
dCTp
(Amersbam
Biosciences/272060)
dGTP
(Amersham
Biosciences/272070)
Primer
(sense/antisense)各種
GoldTM
(Applied Biosystems伽808-0242)
AmpliTaq
lox
PCR
buffer
(PERRIN
)
ELMER侶12387)
DEPC処理水(cALBIOCIiEM/298711)
Crash
< PCR
Master
ice
Mix >
【全て氷上操作】
Table.A-5-3に従い、各試薬を混合しPCRMasterMixを作製する。プラ
イマーは目的に応じて変更する(B-4.
Primer
List参照)。
RT
MasterMix
と同様に、サンプル数が多い場合は1、2サンプル分余分にPCRMasterMix
を作製すること。また、
AmpliTaqGoldT叫ま各サンプルに加える直前に添
加すること。
Table.A-5-3
PCR
Master
(単位:pl)
Mix
12
18
(+2)
(+3)
ll
14
21
28
7
ll
14
21
28
1
7
EE
14
21
28
1
7
1l
14
21
28
1.25
8.75
ll.25
17.5
26.25
35
1.25
8.75
ll.25
17.5
26.25
35
5
35
55
70
105
140
Sample数
1
dÅ∫p(2.5mM)
6(+1)
9(+2)
1
7
dTTP(2.5mM)
1
dCTP(2.5mM)
dGTP(2.5mM)
Primer(sense)(2叫M)
Primer(a山isense)
24
(+4)
(2叫M)
10xPCRbuffer
DEPC処理水
36.25
253.75
398.75
507.5
761.25
1015
AmpliTaqGoldTM
0.25
1.75
2.75
3.5
5.25
7
合計
48
336
528
672
1008
1344
;;t
:車人J、;-,I:人J、:(I:
トノ?I:研J光村
74
Chain
< Polymerase
Reaction
>
【全て氷上操作】
(AmpliTaq
①各サンプルを扱う直前にPCRMasterMix
GoldTM抜き)を
作製する。
② RT法により作製した各RT
しておいたReaction
③
pcRMaster
productから2〃1ずっ採取し、予め用意
Capに移し入れる。
Tubewith
MixにAmpliTaq
GoldTMを加え、vortexによる撹拝とMINI
cENTRIFUGEによる軽い遠心を行う。
㊨ ②で用意したチューブにPCR
Master
vortex
Mixを48J11ずつ加え、
による撹拝とMINICENTR肝UGEにて1分間遠心を行う。
※ GeneAmp㊨
PCR
system
2400
(サーマルサイクラー)を使用する場
合、 pcR中にサンプルが蒸発して組成が変化するのを防ぐため、こ
OIL
(SIGMA/M5904)を
の時点で各サンプルの液面にMINモRAL
50〃1ずつ壁伝いにゆっくりと流し込む。
(参pROGRAMTEMP
CONTROL
ライマーに合ったPCR
SYSTEMにサンプルをセットし、各プ
programを実行する(B-5.
RT/PCR
program
参照)0
⑥作成したpCRproductは-20℃の冷凍庫にて保存する(数ヶ月保存可)0
A15-3.
Gel Electrophoresis
<原理>
核酸やタンパク質などの生体高分子を分析する手段として、電気泳動
法は極めて有効な技術の一つである。電解液を含むアガロ-スやポリア
クリルアミドなどのゲルに電場をかけると、その中を荷電粒子が移動す
る。このとき荷電粒子の大きさ、形、荷電量などの違いによって、粒子
の移動速度に差が生じる。この移動速度の差を利用して様々な分子を分
離する方法が電気泳動法である。
核酸を構成しているヌクレオチドは、塩基とリン酸基が電離して電荷
を帯び易い。 2本鎖DNAの場合、塩基の電荷は相補鎖間の水素結合で互
いに打ち消し合っているため、分子全体としてはリン酸基のマイナス電
荷のみが主となる。また、このリン酸基の個数(荷電)はヌクレオチド
数(DNAの分子量)に比例するため、全てのDNA分子は質量あたり-
75
定の力で引かれることになる。さらに、こうして2重螺旋構造をとって
いるDNAは塩基配列に関わらず同じ線状分子の形をしており、立体構造
は泳動度にほとんど影響を与えない。即ち、唯一泳動度に影響を与える
のが分子の大きさ(長さ)ということになる。
鎖状分子のDNAはゲルマトリックスの網の目の隙間を移動する際、分
子量の大きな長い分子ほどマトリックス-の引っ掛かり(マトリックス
分子との相互作用)が増えるため泳動速度が遅くなる。従って、
DNAの電気泳動の場合、
DNAの塩基配列に関係なく、
2本鎖
DNA鎖の長さの
みに依存した電気泳動パターン(即ち、長いものほど泳動距離が短い)
が得られる。
<注意点>
AGPC法、RT法、pcR法に比べそれほど神経質にならなくても良いが、
ゴム手袋(両手がbetter)と白衣は必ず着用すること。
ぐ意味もあるが、ここではむしろEthidium
RNaseの混入を防
Bromideから身を守るために
あると言っても良い。
@
Ethidium
Bromide
Ethidium
Bromideは核酸のバンド検出に用いられる蛍光試薬であり、
2本鎖DNAの鎖の間に挿入(インターカレーション)されるという
性質上、 DNAの正常な複製や転写を妨げ、突然変異誘起させるとい
う効果から強力な発癌作用と毒性が生まれる。取り扱いには必ずゴム
手袋をはめ、粉末の計量にはマスクを着用すること。また、
Ethidium
Bromideを含んだ泳動用bufferなどの廃液やアガロ-スゲル、廃チッ
プや廃ビンは必ず他のものと区別して集め、適切な処置を施してから
廃棄すること。もしくは業者に委託して廃棄してもらうこと。
また、電気泳動に特有の注意点として、以下の点に気を付けること。
○使用する三角フラスコとメスシリンダーは口にアルミホイルで蓋
をし、
180℃、
8時間以上(overNight)乾熱滅菌を行うこと。
<使用機器・器具・試薬>
・
WideMini-Sub
・
POWERPAC
・
LIBROR
Cell⑧
300
GT
System
(BIORAD/282BR
(BIO
RAD/
170-4485)
11443)
(株式会社島津製作所/AEし200)
:.
・r7_:人J、アニ人芋院 「'、f:研究村
76
電子レンジ
ゲル作成用三角フラスコ(100ml)
メスシリンダー(100ml)
マイクロピペッタ-各種
チップ各種
アルミホイル
サランラップ
(Pechiney
PARAFILM⑧
Plastic
Packaging, Inc.仲M-996)
フライ返し
NuSieve㊥
GTG⑧
Ultra
Agarose
Pure
Agarose
(BMA/50080)
(Invitroge〟155 10-019)
50x
TAE
buffer
(INTERGEN/S4300)
lox
TAE
buffer
(Invitrogen/15558-042)
Etbidium
Blue
123
Bromide
(SIGMA偲-1
5
JuiceTM
10)
(Invitrogen/10816-015)
(Invitrogen/15613-011)
bp DNALadder
DEpC処理水(cALBIOCHEMノ29871
<Agarose
1)
(3% wt/vol)の作製>
Gel
① wideMini-Sub
Cell㊨GT
SystemのGel
casterにゲルトレイをセットし、
水準器を用いてゲルトレイが地面に対し水平になるよう調整する。
② NuSieve㊨
GTG㊨Agaroseとultra
PureAgaroseを秤量し、乾熱滅菌済
みのゲル作成用三角フラスコに入れる(Table.A-5-4)0
※秤量には薬包紙ではなく、乾熱滅菌した三角フラスコの口を包んで
いたアルミホイルを用いること。また、アルミホイルの内側は素手
で触らないこと。
※秤量にはスパチュラを用いないこと。
③ 50xTmbufferを乾熱滅菌済みのメスシリンダーに入れ、
水を所定量加えて50倍に希釈し1x
TAE
buffer
DEPC処理
(&om 50x)を作製する
(Table.A-5-4)
0
Table.A-5-4
Agarose
Gel
UltraPureAgarose(g)
合計(g)
NuSieve⑧GTG⑧Agarose
(蛋)
(3% wt/vol)作製に必要な各試薬の量
0.6
DEPC処理水
(m1)
1.8
1.2
58.
8
合計(m1)
60
50xmEbuffer
1.2
E3]
77
④ ②の三角フラスコに③で調整した1x
mE
buffer
ゲルの塊が出来ないように振り混ぜた後、
(from 50x)を加え、
bufferの蒸発を防ぐため
サランラップでフラスコの口を閉じ、電子レンジを用いて加熱しア
ガロ-スを溶解させる。
※細かい気泡が出なくなり、完全に透明になるまで加熱すること。
※急騰防止のため、様子を見ながら行うこと(目安:l分毎に止める)。
※加熱し過ぎでbufferが多量に蒸発した場合、ゲルの濃度が変わって
しまうため1xTAEbuffer(from50x)を適量加えること。
⑤完全に溶解させたアガロ-ス溶液にEthidiumBromideを6.0〃1加え
てよく振り混ぜた後、ゲルトレイに気泡が出来ないようにゆっくり
と流し入れ、気泡が付着しないようにコ-ムをセットする。
⑥ 10分程度静置しゲルを固めた後、
10倍に希釈して作製した1x
lox
TAE
TAE
buffer
bufferをDEPC処理水で
(from lox)にてゲル表面を浸
し、冷蔵庫に10分程度入れて完全に固める。
< Gel Electrophoresis
>
①水準器を用いて、地面に対して水平になるように調整したWide
Mini-Sub
600ml、
Cell㊨
GT
Systemの泳動槽に、
EthidiumBromideを20〃
1Ⅹ TAB
buffer
(from lox)を
1加える。
②完全に固まったゲルからコ-ムを慎重に抜き取り、サンプル側が陰
極となるようゲルトレイごと泳動槽に移し入れる。
※ ゲル本体及び穴が崩れた場合は作り直すこと。
③ pMILM⑧を20cm程度切り出し、その上にBlueJuiceTMを3〃1ず
つ滴下する。
※
Blue
JuiceTMはすぐに乾燥してしまうため、
1度に全サンプル分を滴
下しない方が良い。
④
123
(3pl)は、
bpDNALadder
BlueJuiceTM
(3〃1)及び1xTAEbuffer
(from50x) (3〟1)と混合し、ピペッティング後にゲルの所定の穴に
静かに流し込む。
⑤各pcRproductから10〃1ずつサンプルを採取し、
BlueJuiceTM
(3〟
1)に加えた後ピペッティングを行い、ゲルの所定の穴に静かに流し
込む。
⑥
positive
Controlを10〃1採取し、所定の位置のBlue
JuiceTM
(3〃1)
に加えた後ピペッティングを行い、ゲルの所定の穴に静かに流し込
む。
※混合後の溶液(13〃1)は残さず吸い取り、流し込むこと。
I.車)(
(ijr・
J寛村・
Jt;':八′、デ:院「 ′、i,[:
78
⑦電流がゲル内を真直ぐに流れるようにゲルトレイの位置を調整する。
⑧電極を取り付けpOWERPAC
300に接続した後、
80V
80mA70分の
条件で泳動を開始する。
A-5-4.
PCR反応生成物の確認と定量化
<原理>
核酸は280nm付近の波長に吸収ピークがある。この核酸のUV光を吸
収する性質は、対象となる2本鎖DNAの内部にEthidiumBromide分子を
挿入(インターカレート)することでPCR反応生成物の確認に応用する
ことが出来る。即ち、核酸(ここでは2本鎖DNA)に吸収された280nm
のUV光がEthidium
Bromide分子にエネルギー転移されると、励起され
て590nlnの蛍光を放射する。
Ethidium
で、この300nmの波長をあまり含まず、
uv光でEthidium
Ethidium
Bromide本来の吸収は300nmなの
280nm付近の波長を豊富に含む
Bromide染色を行ったゲルを照らすと、遊離した
Bromide分子は励起されず、インターカレー卜した分子のみが
DNAからのエネルギー転移により励起されて蛍光を発する。つまり、ゲ
ル中で2本鎖DNAが存在する部分のみが光るということになる。この蛍
光強度は概ねインターカレー卜したEtbidium
Bromide分子の個数に比例
し、また、インターカレーションは2本鎖DNAの量(長さ×本数)に比
例して起こるため、蛍光強度を測定することでゲル中のDNA量を推定す
ることが可能になる。但し、測定する2本鎖DNAが全て十分量のEthidium
Bromideと結合し、飽和していなければ、その定量は意味を成さないので
注意しなくてはならない。
<内部標準>
内部標準とは、それぞれ検出したサンプル量を補正するために必要と
なる基準のことである。定量的pcR一般において一義的に明らかにされ
ることは、反応系に加えたサンプルに含まれる目的DNAの相対量であり、
絶対量ではない。その絶対量を知るためには、予め絶対量の分かってい
るDNAで標準サンプルを調整してPCRを行い、目的DNAと標準サンプ
ルとの相対量を比較する必要がある。この標準となるものを内部標準と
(グリセルアルデヒト3-リン酸脱水素酵
いう。本研究で用いたGAPDH
素)はbousekeepinggeneの一種であり、組織によって発現量に差がない
ため内部標準として用いることが出来る。また、GAPDHの他にもα-actin
やβ-actinなども内部標準としてよく用いられる。
79
-補正の具体例2つのサンプルA、
える。
(目的物)、内部標準遺伝子Nを考
Bと遺伝子Ⅹ
AとBそれぞれにおけるⅩの発現量を比較する場合
i)遺伝子Ⅹの定量結果A:ち-a:bだけではA中のⅩはB中のⅩの
〟b倍多い(もしくは少ない)量が入っているという「相対量」しか
分からない。
立)ところが、絶対量が分かっている内部標準遺伝子Nの定量結果A:B
nを知るこ
-m:nからA中のNとB中のNそれぞれの絶対量m、
とが出来る。また、
A中のNとB中のNそれぞれの相対量(m/n倍
多い、少ない)も知ることが出来る。
近)そこで、
i)で分かったxの相対量を、
ii)で分かったNの絶対量
で補正すればⅩの発現量の絶対量〟mとb/nを求めることが出来る
(Table.A-5-5)
。
Table.A-5-5
目的遺伝子の相対量と絶対量
A
B
a
b
目的遺伝子Ⅹの相対値
Nで補正したⅩの相対値
相対
量
内部標準遺伝子Nの相
m
n
〟m
b/n
対値
量
絶対
量
<注意点>
ここでもEthidium
Bromide汚染の危険性があるため、その扱いには十
分注意すること。また、
pCR反応生成物の確認と定量化に特有の注意点
として、以下の点に気を付けること。
○ゲルは折れやすいため、無理な力を加えないこと。
○ゲルをイメージアナライザーに付属のプレートに移す際は、隙間に
気泡を入れないこと。
○フロッピーディスクに画像データ(jpeg型式)を記録する際は、自
動的にファイル名が連番の番号のみとなるため、大量に撮影する場
合はどの画像がどの結果なのか確認できるようにしておくこと。
A.
I;I;t
)(一?':
,T7:人J、デ:
「
J、i::桝'')YJ杓
80
<使用機器・器具・試薬>
・イメージアナライザーー式(TOYOBO/FAS-Ⅲ)
・
Analyzer
Gel-Pro@
Cybernetics⑧/50N3
Version
3.1
for
(
WindowsTM
Media
1 000- 1 1 85 1 )
・パーソナルコンピュータ
(Windows⑧フォーマットのもの)
・フロッピーディスク
・サランラップ
<pcR反応生成物の確認>
①電気泳動終了後、ゲルをイメージアナライザーに付属のプレートに
移してセットした後、モニタに写ったゲルの位置、被写体の大きさ
を調整し、カメラのフォーカスを合わせる。
②uvトランスイルミネ一夕-より
UV光を照射し、バンドの確認を
行う。
③鮮明なバンドの確認が出来た場合、露光時間を変えて数パターンの
写真を撮影し、付属のドライブよりフロッピーディスクに画像デー
タ(jpeg型式)を記録する.
④ positive Controlを含め、バンドが確認できなかった場合は、作製し
たゲルや電気泳動操作に問題があったと考えもう一度やり直す。
⑤記録した画像データをパソコンに移し、画像解析ソフト
Analyzer
Version 3.1
Gel-Pro⑧
forWindowsTMを用いてバンドの解析を行う.
<データの扱い・処理>
画像解析ソフトGel-Pro㊨Analyzer
Version 3.1 for WindowsTMを用いて以
下の2点を解析する。尚、解析操作はソフトに付属の取扱説明書に従っ
て行う。
i
)サイズマーカーと比較し、
PCR反応生成物のSize
(bp)を解析する。
近)各泳動レーン内の目的のバンドを選定し、バンドの輝度(max.oD)
を測定する。
得られた各データ値は内部標準のデータ値で除し、
controlを1として
補正する。
81
A-伝.ウェスタンプロット法
◎測定の原理
・BCA法
蛋白はアルカリ溶液中で2価の銅イオンに作用し、
1価の銅を生成させます。
(BicinchoninicAcid)分子と錯体を形成し
この1価の銅イオンが2分子のBCA
紫紅色を発します。その際の吸光度を測定することでタンパク質濃度を求め
る
・
SDS-PAGE
SDSは水溶性タンパク質1g当たり約1.4g結合してSDS一夕ンパク質複合体を
形成します。タンパク質分子自身が持つ電荷はSDSとの結合によってほとん
ど打ち消されている。また、還元剤である2-メルカプトエタノールによって
タンパク質のS-S結合(ジスルフィド結合)が切断され、
SDSが結合したタ
ンパク質の形状はほぼ直鎖状になっているため、タンパク質分子の本来の形
状は移動速度に影響を及ぼさない。これらのことから、ゲル内でのタンパク
質の移動のしやすさは分子の大きさとそれに比例した負電荷の量にのみ左右
され、分子量に従って分離することができるo
ウェスタンブロツテ
漉紙の間にゲル、膜を挟み、電圧をかけることでタンパク質を電気的にゲル
から膜-転写する方法。
・化学発光検出
化学発光は化学反応によって生じたエネルギーが光に変換されて起こるもの
で、その中でもアルカリ条件下におけるHRPと過酸化水素によって触媒され
るルミノール酸化反応がある。ルミノールは酸化されると直ちに励起状態と
なり、基底状態-と減衰する過程で光を放ちます。
ECL反応はフェノール環
を持つ化合物などのエンハンサーの存在下でHRPによってルミノールが酸化
されたときに起こる。
-.
i;JJL(: 卜′、デ・'研''JtJi村
ET,:人下人ノi三:
82
A-6-1.細胞からタンパク質の抽出
<使用機器・器具・試薬>
6穴プレート(住友ベークライト/NS80060)
・
・高速冷却遠心機(HIMAC
・
PBS
CR1
(SIGMAの8537)
IiEPES
(Wako/340101371)
・
protease
inhibitor
・
phosphatase
・
5B,BIO-RAD)
(SIGMAJP8340-1ML)
inhibitor
<操作>
① connuentの75cm2
(SIGMAn5726-1ML)
naskの培地を吸引し、PBSIOmlで十分に洗浄する。
※氷上で行う
②細胞溶解液(o.1%
TritonX-100を含む20mM
HEPES-NaOH
,
pH7.2)をフ
ラスコに1.Oml加える。
③
protease
inhibitor、 pbosphatase
inhibitorをそれぞれ5
〟
1加える。
④ 25-37℃で5-20分間、軽く振返しながらインキュベ-卜し、細胞膜を
溶解させる。
⑤セルスクレーパーで細胞を剥離し(または強くピペッティング)、
1.5ml
tubeに移す。
⑥冷却遠心機で15,000rpm、
30分間、遠心する。
⑦上澄み液をアシストチューブに移す。
※抽出したタンパク質は冷凍保存(-ュo℃)可能
83
A-6-2.タンパク質の定量(BCA法)
<使用機器・器具・試薬>
spectrophotometer
・
(SmartSpecTMplus,BIO-RAD)
・ビシンコニン酸(ナカライテスク/04841-54)
・酒石酸ナトリウム(Wako/197-13975)
・
Na2CO3
・
NaOH
・
NaHCO3
(Wako/196-01595)
(Wako
/196-05635)
(Wako/191-01305)
・硫酸銅(Ⅲ)5水和物(wako/033-04415)
<操作>
① A溶液、
B溶液をそれぞれ作製する。
A溶液(数ヶ月室温保存l
・
ビシンコニン酸
1g
酒石酸ナトリウム
0.16g
Na2CO3
2g
100mlにメスアップ後NaOH
NaOH
0.4g
pHll.25にする。
NaHCO3
0.95g
DW
適量
を加え、
・
B溶滅(室温保存)
CuSO4
・
5H20
4g
DW
IOOml
②スタンダード(1mg/mA
1mg/m]BSA
EZZZZ]
BSA)を作製し、下表のように加える。
DVV
言式馬兵曹1本当たり
EZZZZ]
のBSA量(〟l)
0
10
0
2
8
2
4
【ヨ
4
6
4
6
8
2
8
10
0
10
:.
)( ′、㌢:
)、こ`、il二院「 J';::研′先約
LTT_;
84
③ A溶液とB溶液を50:
1で混合する。
④サンプルとスタンダードをそれぞれ10〃1ずつアシストチューブにと
り(各サンプル2本ずつ)、
③で作製した溶液を10011ずつ加える。
⑤ボルテックスで授拝する。
⑥ 37℃、
30分で静置する。
⑦ OD562を測定する。
⑧タンパク質濃度を横軸、吸光度を縦軸にとり、検量線を作り、総タン
パク質量を測定する。
* spectrophotometer
(SmartSpecTMplus.
BIO-M)操作方法
1.電源を入れる
2.入を押す
3.enterを押す(波長の読み取り1)
4.波長を562nmに設定する
5.enterを押す(バックグラウンド読み取りをひきますかNo)
6. select
-
enterを押す(繰り返して読むサンプルはありますかYES)
7.
enterを押す(試料の反復番号3)
8.セルにBla止(PBSのみ)を入れ、
Read
Blankを押す
9.
-→を押す
10.試料を戻して、セルを脱イオン水で洗浄し、脱イオン水を取り除く
ll.セルに試料を入れ、
ReadSampleを3回押す
12.試料を戻して、セルを脱イオン水で洗浄し、脱イオン水を取り除く
13.
10-11を繰り返す
85
A-6-3.
SDS-PAGEによる蛋白質の分離
<使用機器・器具・試薬>
(ATTO/MAB110、
・ゲル板(前面ガラス板、ミミ付ガラス板)
MB-00)
・シリコンガスケット(ATTO/MS-01)
・プラスチック製クリップ
・コ-ム(ATTOAMIO-12)
・電気泳動槽(AE-6410型,ArTO)
・電源装置(クロスパワー500,ÅrTO)
Tris(トリスヒドリキシメチルアミノメタン)
・
・
・
(Wako/199-07141)
SDS(ドデシル硫酸ナトリウム)
(Wako
HCl
(Wako/207-06275)
/080-08065)
・アクリルアミド(wako/016-00765)
・
Bis(N.N'-メチレンビスアクリルアミド)
・
APS(過硫酸アンモニウム)
(wako/138-06032)
(ナカライテスク/02602-15)
・グリシン(Wako/077-00735)
・
・
(Wako/021-02911)
BPB(プロムフェノールブルー)
2-メルカプトエタノール(Wako/135-07522)
・グリセロ-ル(Wako/079-00614)
TEMED(N,N,N',N'-テトラメチルエチレンジア
ミ
・
ン)
(SIGMA/T928ト25ML)
・分子量マーカー(BIO-W/161-0318)
<準備>
1)電気泳動用試薬の調製
I
(pⅡ8.8),0.4%SDS
I.5MTris-IICl
Tris54.5g、
-
4℃で保存
SDSl.2gを脱イオン水に溶かし、
5N
HCl(約17ml)を加え
てpH8.8に合わせ、脱イオン水で300mlにメスアップする。
・
o.5M
Tris-ⅡCl
Tris6.Og、
(pII6.S),0.4%SDS
-
4℃で保存
SDSO.4gを脱イオン水に溶かし、
5N
HCl(約9ml)を加え
てpH6.8に合わせ、脱イオン水で100mlにメスアップする。
I
o.5M廿is-HCl
(pⅡ6.S)
4℃で保存
-
5N HCl(約5ml)を加えてpH6.8に
Tris3.Ogを脱イオン水に溶かし、
合わせ、脱イオン水で50mlにメスアップする。
4℃で保存
・30%アクリルアミド溶液アクリルアミド58g、
Bis2.Ogを脱イオン水に溶かし、脱イオン水
で200mlにメスアップする。
:_車人乍人ノ、i:二院
l-.L、)1f二研駕村
86
o.4%SDS
・
-室温で保存
sDSO.2gに脱イオン水50mlを加えて溶解する。
-当日作製
・10%APS
APS
0.1gに脱イオン水1mlを加えて溶解する。
・泳動バッファーー→室温で保存
Tris3.Og,グリシン14.4gを脱イオン水に溶かし、脱イオン水で
1000m】にメスアップするoその後、
・サンプルバッファー0,5MTris-HCl
sDSl.Ogを加え、溶解させるo
4℃で保存
(pH6.8)2ml、 0.4%
SDS4ml、
2-メルカプトエタノー
ル1.2ml、グリセロ-ル2mi、脱イオン水o.8ml、
0.1%BPB数滴を
混ぜる。
・ゲル保存用バッファー(mming
1.5M
Tris-HCl(PHS.8) 0.5ml、
gelを保存する際にゲル上に重層する)
0.4%SDS
20LL
l、脱イオン水1.4Smlを
混ぜる。
■I1■■■■Ll■l■L
runmnggeI
<換作>
Ⅰゲル板の組み立て
(丑前面ガラス板とミミ付ガラス板、シリコンガスケットをエタノールで拭
くo
野
・
/■■L
庭さ▲
:l=:_:・
・
前面ガラス板
ミミ付ガラス板
シリコンガスケット
87
コ-ム
クリップ
② ミミ付ガラス板にシリコンガスケットをセットし、前面ガラス板をのせ
て、クリップでとめる。
云二「
・・_i
I
③コ-ム(comb)を差し込み、コ-ムの下底から5mmのところに目印を入
れる。
/(
.
LIT.:
′
J::l」
`、j::ド:.i r,て-阿1PfH・:r
88
II
running
gel、
stacking
gelの作製
Tabale.A-5-1アクリルアミドゲル作成
アケノルアミド濃度(%)
REAGENT
RunnnjnggeL
5%
30-200kD∈l
分画分子量範囲
Stackinggel
7.5%
10%
12.5%
40-200kDa
20-130kDa
14-80kDa
2.Oml
3.Oml
4.Oml
5.Omf
3.Oml
3.Oml
3.Oml
3.Oml
4.5%
30%
0.9ml
アクリルアミド溶さ夜
1.5MTris-
HCl(pH8.8).
0.4%SDS
0.5MTris-
HCl(pH6.8),
1.5ml
0.4%SDS
APS
120JJl
120JJl
120JJL
120〃l
60fLl
TEMED
12FLL
12〟l
12JJl
12FJl
6FLl
脱イオン水
TotalVoーume
※
7.Oml
6.Oml
5.Oml
4.Oml
3.6ml
12.Oml
12.Oml
12.Oml
12.Oml
6.Oml
目的とするタンパク質の分子量に対応して、ゲル濃度を決定する0
※表はゲル1枚分の量を示している。
※アクリルアミドのモノマーは神経毒であるので、試薬を扱う時は手袋、マ
スクを着用する。
①running
gel溶液、 stacking gel溶液を上の表に従って作成するo
TEMEDはこの時点では加えない)
②ruminggel溶液をアスビレータ-で脱気する.
③脱気後、
(但し、
TEMED12〃1を加え、軽く混ぜる。
※微量なのでピペット操作は要注意。
④素早く、
2)で準備したゲル板の目印のところまでmnninggel溶液を入れ
る。
※ フラスコから直接注ぎ込む。
⑤ ピペットで脱イオン水をゲル液面上に重層する。
⑥ゲル化するまで静置する。
(5-10分程度)
※アクリルアミドは20℃以下では重合しにくいため、室温に注意。
(特に
冬期は注意)
※ゲルと水の界面ができていればゲル化しているので,水を捨ててキムワ
イプで軽く拭く。
89
⑦ゲル化させている間にstackinggel溶液をアスビレータ-で脱気する。
⑧脱気後、
TEMED6〃1を加え、軽く混ぜる。
※微量なのでピペット操作は要注意。
⑨素早く、ミミ付ガラス板の切り欠きの少し下までstacki喝gel溶液を入
れる。
⑩コ-ムを気泡が入らないように差し込み、約30分間静置してゲル化さ
せる。
※ コ-ムを両手で持ち、水平に差し込む。
Ⅲ各ウェルにローディングする試料溶液の調製
①各ウェルに加えるタンパク量(100-200〃g)に相当する試料の体積を
BCA法により測定した濃度から算出する。
② (丑で算出してタンパク量とサンプルバッファーを1:1で混合する.
③ボルテックスで獲拝する。
④ 100℃の熱湯で3分間ボイルする。
⑤ボルテックスで摸拝する。
Ⅳ電気泳動
①電気泳動槽に底から(陽極側)泳動バッファーを入れる。
②ゲル化したらコ-ムをそっと抜き取る。
③クリップとシリコンガスケットを外し、ゲル板を締め付け具に固定し、
電気泳動槽に入れる。
④ゲルの上部(陰極側)が十分に浸るぐらい電解槽液を入れる。
⑤サンプルをピペットでウェル内にゆっくり注入する。
※ガラスにマジックでサンプル番号を書くと見やすい。
※分子量マーカーはサンプルバッファーを加えずに、分子量マーカー5〃1
のみ注入。
⑥電極端末を接続し電源を入れ、ゲル1枚のときは15-20mAの定電流で、
ゲル2枚のときは30mAの定電流で泳動する。
※ BPB線がゲル下端から5mmの位置にくるまで泳動する。
A-6-4.ゲルの染色(cBB染色)
<使用機器・器具・試薬>
・メタノール(Wako/134-01833)
・酢酸(wako/012100245)
・cBB-R250(クマシープリリアントブルー)
(ナカライテスク/09408-52)
トノ、i::研′')t手:t
・-.求人′';・':人′、デ:育ちこ
90
<準備>
・前固定液-当日作製
メタノール20ml、酢酸7.5ml、脱イオン水72.5ml
・染色液-室温で保存&再使用
CBB-R2502.5g、メタノール500ml、酢酸50ml、脱イオン水450ml
・脱色液-室温で保存&再使用
メタノール50ml、酢酸70ml、脱イオン水880ml
<操作>
(丑容器にゲル全体が浸るぐらいの前固定液を入れておく.
②電気泳動の終わったゲルを前固定液に入れる。
③室温で30分間振塗する。
④前固定液を捨て、染色液を入れる。
⑤室温で30分間振返する。
⑥染色液をビンに戻し、脱色液を少量入れる。
⑦容器を軽く揺すり、染色享夜をすすぎ,脱色液を捨てるo
⑧脱色液をたっぷり入れて、キムワイプを2,3枚脱色液に浸けこむ。
※脱色の際にキムワイプを適宜交換することで、脱色を早くすること
が出来る。
⑨室温で数時間-一晩振塗する.
A-伝-5.ゲルから膜-の転写(ウエスタンブロツティング)
<使用機器・器具・試薬>
・
(ArTO/2392390)
pvDF膜(ポリフッ化ビニリデンメンプレン)
・ブロツティング漉紙(ArTO/2392393)
・ブロツティング装置
Tris(トリスヒドリキシメチルアミノメタン)
(Wako/207106275)
・
・メタノール(Wako/134-01833)
6-アミノカプロン酸(wako/018-09642)
・
<準備>
・
A溶液(o.3%MTri$
5%メタノール溶硬)
-
4℃で保存
Tris3.6342gを脱イオン水50miに溶かし、5mlのメタノールを加えて、
最後に脱イオン水を加えて計100mlとする。
91
B溶液(25mMTris
・
5%メタノール溶液)
-
4℃で保存
TrisO.3029gを脱イオン水50mlに溶かし、5mlのメタノールを加えて、
最後に脱イオン水を加えて計100mlとする。
-
40mM6-アミノカブロン酸
・c溶液(25mMTris
4℃保存
Tris 0.3029g、
40mM
5%メタノール溶硬)
6-アミノカブロン酸o.5247gを脱イオン水50ml
に溶かし、 5mlのメタノールを加えて、最後に脱イオン水を加えて
計100mlとする。
<操作>
ウエスタンブロツティング
①電気泳動終了前に、
PVDF膜をmnningelと同じ大きさに切り、メタ
ノールに20秒ほど浸し、次にB溶液に浸し30分ほど振塗させる。
②さらに、電気泳動終了前に、ブロツティング漉紙6枚をmnning
gel
と同じ大きさに切り、A溶液に2枚、B溶液に1枚、c溶液に3枚浸す。
③電気泳動終了後、ガラス板の隙間にスパチュラを差し込み、てこの
原理でガラス板をはがす
④ stackinggelを取り除き、 mnninggel部分のみにして、
B溶液に浸す。
⑤準備してあったブロツティング漉紙で、まずA溶液に浸した漉糸氏2
枚をブロツティング装置にのせる。さらに、
B溶液に浸した漉紙1枚、
pvDF膜、ゲル、
C溶液に浸した漉紙3枚の順に上-重ねる。
⑥グローブをはめ、手のひらで全体を押しつぶすように気泡を抜き、
膜とゲルを密着させる。
⑦一番上の漉紙にC溶液をかけ、陰極板をセットし、リード線をつな
ぐ。
⑧ゲルの面積1cm2ぁたり2mAの定電流で約40分間ブロツティングす
る。
A-5-6.抗体処理
<使用機器・器具・試薬>
・スキムミルク
・
・
・
・
・
PBS
(SIGMA/D8537)
pBS粉末(日水製薬/o5913)
TritonX-100
Monoclonal
Rabbit
(SIGMA/T8787-50ML)
Mouse
polyclonal
Anti-Human
to
beta Actin
Smooth
Muscle
Actin
(Dako/MO85
1)
(abcam/ab 16039-500)
:▲帝人`、;:二人Jl;-I:院LJl;I:研′発車斗
92
・
HRP-Goat
anti-Rabbit
lgG
(H+L)
(Zymed/62-6120)
I
HRP-Goat
anti-Mouse
IgG
(H+L)
(Zymed/6216520)
一次抗体反応
<準備>
・PBS
pBS粉末9.6gを脱イオン水に溶解して全量を1000mlにする。
・
0.1%PBST
PBS
19にTritonl.Omlを加える。
・ブロッキング液(1%スキムミルク/0.1%PBST)
pBST50mlにスキムミルク0.5gを加える。
・一次抗体
ブロッキング液1.Omlに一次抗体2.OJ11を加え500倍希釈する。
<操作>
①ブロッキング液に膜を浸し、
② pBSTで10分毎に3回洗浄。
overnightブロッキングする。
(4℃)
(室温)
※ ゆるやかに振塗させる。
③ケースにパラフイルムをセットし、その上にPVDF膜をのせ、一次
抗体溶液を添加して室温下で1時間静置させる。
※抗体溶液が移動しないように水平にセットする。
二次抗体反応
<準備>
・二次抗体
ブロッキング液1.OmlにHRP標識二次抗体(HRP-Goat
を1.0〃1加え、
anti-Rabbit
lgG)
1000倍希釈する。
<操作>
(∋ pBSTで10分毎に3回洗浄。
②ケースにパラフイルムをセットし、その上にPVDF膜をのせ、二吹
抗体溶液を添加して室温下で1時間静置させる。
※抗体溶液が移動しないように水平にセットする。
③ (丑の操作を行う.
93
A-6-7.バンドの定量
<使用機器・器具・試薬>
・
・
Variable Model
ECL
Imager(Typhoon 8600,Amersham)
(Enhanced CbemiLuminescence) (Amersham/RPN2
1 09)
1液と2液を500〃ずつとる。測定直前に混ぜる。
※アルミで遮光、氷上
※1液と2液は等量混ぜ、
o.125ml/cm2を目安にECLの量を決め
る。
<操作>
(D洗浄した膜をラップ上にのせる。
② ECLの1液、
2液を混ぜ、膜上に添加する。
※ 1分間静置させる。
③新しいラップ上に膜を移し、ラップで包み込む。
※
しわがつかないように包む
④ Typhoonで検出を行う。
⑤ Imagequantでタンパクの定量を行う。
※
Typhoonの操作方法
1.テーブルタップ・コンピュータ・Typhoon本体の電源スイッチをONにす
る。
2.スタートアップスクリーンが表示されたら、
Ctrl,Alt,Deleteキーを同時に押
す。
3.Welcomeウインドウが表示されたら、
oKをクリックする。
(パスワードな
し)
4.サンプルをガラスプレートの上にセットする。
(左下を基準にセットする)
5.デスクトップ上の「Typhoon3.0」のアイコンをクリックして起動させるo
6.スキャンエリアを指定する。
(スキャンの開始位置の座標をマウスの左ボタ
ンでクリックしそのままスキャンするエリアをドラッグする)
7.スキャンモードを「chemiluminescence」にする。
8.
setupをクリックし、
定する。
9.
sensitivityを「high」、
PMTボルテージを「950」に設
Orientationでサンプルの表裏と方向を決定する。
10.ピクセルサイズを指定する。
(50〃1,100〃1,200〃1から選択する)
ll. Im喝e
(〕uantに指定する。
Analysisの項目をImage
12.SCANボタンをクリックして検出を行う。(sCANボタンをクリックすると
:,車人ノ、;・r:
I(、;・':院
f1.
1、;'・'L7L)卜先約
94
SaveAsウインドウが表示されるので、サンプルのイメージデータに名前
を付けて保存する)
13.スキャン終了後、スキャンした画像が表示される。
※ Imagequantによる定量方法
1.イメージの最適化
(1)ツールバーのGrey/Color/Adjustボタンをクリックする。
(2)Highスライダー(上側)をドラッグして、プレビューイメージ内のバンド
が赤色に表示され始めたら、マウスボタンを放します。
(3)Lowスライダー(下側)を打ラッグして、プレビューイメージ内のバック
グラウンドが紫色に表示され始めたら、マウスボタンを放します0
(4)OKをクリックするo
用され、
(イメージウインドウの表示イメージに変更内容が適
Grey/Color/Adjustウインドウが閉じる)
2.レクタングルの作成
(1)ツールバーのRectangleボタンをクリックする。
(2)目的のバンドにポインターを当て、バンドがレクタングルで完全に囲ま
れるまで、ポインターをドラッグします。
(3)Selectボタンをクリックする。
(4)作成したレクタングルの内部中央にポインターを合わせ、マウスボタン
を押したまま、キーボードのⅤキーを押す。
(5)マウスボタンを押したまま、次のレーンのバンドがレクタングルで囲ま
れるように移動させる。
(6)この操作を繰り返し,各レーンのバンドにレクタングルを作成する。
3.ボリューム解析の評価
(1) Selectボタンが選択されていることを確認する。
(2)レクタングルの1つをクリックする。
(3)ツールバー(下)のⅦ1umeReviewボタンをクリックすると解析評価が表
示される。
95
A-7.三次元遊走モデルの作成
A17-1.
Type
l
collagen
gel溶液の作成
<使用機器・器具・試薬>
(MILLIPORE/Cat.No.SLGVO25LS/孔径o.22ドm)
Millex-GV
・
strivex
・
GS
(MILLIPORE/Cat.
No.SVGSBIOIO/孔径o.22
pm)
15ml遠心管(住友ベークライト/MS-56150)
・
50ml遠心管(住友ベークライト/MS-56500)
I -A
cellmatrixType
(新田ゼラチン/637-00653)
・
・
HEPES
・
2-[4-(2-Hydroxyethyl)-トpiperazinyl]ethanesulfbnic
acid
(wako/340-01371)
(Wako/038-10531)
NaHCO3
・
1NNaOH(196-05635/Ⅵねko)
・
1
1NHC
・
(080-08065Nako)
10倍濃縮培地(M199倍地)
・
(新田ゼラチン/02-069-010)
<準備>
##
A.3.Omg/ml
pH3.0
Cellmatrix
Type
I
-A
試薬B.10倍濃縮培地
試薬c.再構成用緩衝溶液
0.05N
させ、
2.2g、
NaOH溶液100mlに対してNaHCO3
HEPES4.77gを溶解
0.22〟mフィルターで漉過滅菌を行った。
※大量に貯蔵しておくと、
NaHCO3からCO2が抜けていくため、
5ml
ずつ分注して密封保存しておく。
<操作>
しっかりと低温状態を保つため、全て氷上操作で行う。またゲル濃痩
によって各よう液の混合量が異なるためTable.A-6-1に従うこと。
①
A、
B、
C液を8:1:1となるように50ml遠心管内で混合する。この
際A手夜とB液を混合した後にあわ立てないようによく撹拝し、さ
らにC液を加えよく混ぜる。
※色を見ながらpH7.4になるように、緩衝溶液の量で調節する。あ
るいは1N
HClやNaOHで調節を行う。
5〃1前後を加えながら微調
:.有人J'i・':八一、デ:院 L /、i::
Li)千
'先約
96
整するとよい。
※あらかじめpH7.3、
7.4、
7.5の標識標本を水:10倍濃縮培地:緩衝溶
液が8:1:1となるようにして作っておくこと。
②
M199培地を加え、ゆっくりと振り混ぜる。
それぞれのゲル濃度の溶i夜を1.8ml作るとすると、それぞれの溶液の割
合は以下の表のようになる。
Type
Table.A-6-1
I
collagen
gel作成
ゲル濃度(mg/m1)
E3)
2.0
1.5
1.0
0.5
CellmatriX
1200
900
600
300
濃縮培地199(〃1)
150
112.5
75
37.5
緩衝溶液(〟1)
150
112.5
75
37.5
M199培地(〃1)
300
675
1050
1425
Ⅰ-A(〃1)
A-7-2.
Fibronectin
in Type
I
collagen
gelまたはBSA
in type
l
collagen
gel
溶液の作成
<使用機器・器具・試薬>
・
・
・
・
・
・
・
・
・
・
・
・
Millex-GV
(MILLIPORE/Cat.No.SLGVO25LS/孔径o.22
pm)
strivex
(MILLIPORE/Cat.
pm)
GS
No.SVGSBIOIO/孔径o.22
15ml遠心管(住友ベークライト株式会社/cat.No.MS-56150)
50ml遠心管(住友ベークライト株式会社/cat.No.MS-56500)
(新田ゼラチン/637-00653)
I -A
cellmatrixType
HEPES
2-【4-(2-Hydroxyetbyl)-1-piperazinyl]etbanesulfbnic
acid
(wako/340-01371 )
(Wako/191-01305)
NaHCO3
1NNaOH(196-05635Nako)
1NHC
1
(080-08065Nako)
10倍濃縮培地(M199倍地)
BSA
(Wako/019-07494)
Fibronectin
(新田ゼラチン/02-069-010)
(三光純薬/341635)
※原液濃度1g/ml
⇒
M199培地で50pg/mlに希釈して分注後、冷
凍庫にて-20℃で保存。解凍する際にはゲル化してしまうため振
り混ぜないこと。
97
<準備>
試薬A.3.Omg/ml
Cellmatrix
pH3.0
TypeトA
試薬B.10倍濃縮培地
試薬c.再構成用緩衝溶手夜
2.2g、
NaOH溶液100mlに対してNaHCO3
o.o5N
させ、
HEPES4.77gを溶解
0.22〃mフィルターで漉過滅菌を行ったo
※大量に貯蔵しておくと、
5ml
NaHCO3からCO2が抜けていくため、
ずつ分注して密封保存しておく。
<操作>
しっかりと低温状態を保つため、全て氷上操作で行う。またゲル濃痩
によって各よう液の混合量が異なるためTable.A-6-2に従うこと。
(丑A、
B、
C液を8:1:1となるように遠心管内で混合するo
この際A液
と B液を混合した後にあわ立てないようによく撹拝し、さらにC
液を加えよく混ぜる。
※色を見ながらpH7.4になるように、緩衝溶液の量で調節する。あ
るいは1N
HClやNaOHで調節を行うo
5FLl前後を加えながら微調
整するとよい。
※あらかじめpH7.3、
7.4、
7.5の標識標本を水:10倍濃縮培地:緩衝溶
液が8:1:1となるようにして作っておくこと。
② FNまたはBSA及びM199培地を加え、撹拝する。
それぞれのゲル濃度の溶液をl.8ml作るとすると、それぞれの溶液の割
合は以下の表のようになる。
TableA-6-2
(〃1)
2.0
Type
FN及びBSAin
I
collagengel作成
ゲル濃度(mg/m1)
1.5
1.0
0.5
CellmatriX
l200
900
600
300
150
112.5
75
37.5
150
l12.5
75
37.5
300
315
690
1065
360
360
Ⅰ_A
濃縮培地M199
緩衝溶液
M199培地
FNまたはBSA
360
(〃1)
I;I;(三 トノi・':岬J!lt手ごL
`′帝人′';'ニセ′、jf:
98
※FNまたはBSA有りの場合(ゲル作成時はFN濃度10FLg/ml
散後は5pg/ml、
A-713.
ⅡPMC
⇒
BSA濃度7.5FLg/ml
⇒
拡
1.5Jlg/ml)
βl誘導によるEMTの誘導
-のTGF-
<使用機器・器具・試薬>
6穴プレート(住友ベークライト/NS80060)
TGトβ 1 (SIGMA/T7039-2-UG)
・
・
・
M199
・
FちS
(SIGMA/F9423)
(SIGMA/F9423)
<操作>
① 6穴プレートにHPMCを播種し、subconfluentになるまで培養する。,
② o.1%FBS仙I199で24時間growth
arrestする。
③
6
穴プレートの培地を吸引し、
3
1/0.1%FBSバM199を2ml/we11ずっ加え、
穴分に10喝/mITGF-β
48h培養するo
A-7-4.トランスウェルを用いたⅡpMCの問質-の遊走モデルの作成
<使用機器・器具>
・ミリセルPCF
(膜孔12〃m)
(MmLIPORE/PIXPO1250)
24穴プレート(住友ベークライト/MS-80240)
・
APPENDIXA-5-1で作成した混合用液
・
APPENDIXA-5-2で作成した混合用液
・
(SIGMA仙蛇154)
M199
・
・
・
FBS
(SIGMA/F9423)
PBS
(SIGMAD8537)
TRYPSrN-EDTASOLUTION
・
(lox) (SIGMA/T4174)
・血球計算盤
<換作>
①あらかじめ膜を300〟1のM199で浸しておく。
② APPENDIXA-5-1またはAPPENDIXA-5-2で作成した1.5mg/mlコラ
ーゲンゲル200plを24穴プレートlowerに加え、均一になるよう
に整える。
※ピペットを押し切ると空気が入るため、ゲルに空気が入らないよ
うに注意する事。
③ 37℃で30分間インキュべ-トする。
99
④ ②で作成したゲル上にAPPENDIXA-5-1で作成したo.5mg/mlコラー
ゲンゲル200〃1を積層し、均一になるように整える。
⑤ (Dの膜から培地を抜き取り、コラーゲンゲル上-乗せる。
※膜とゲルの間に空気が入らないように、ゆっくりと乗せる。
⑥ o.1%FBS/M199
または72h、
300〃1を膜に加え、
37℃でFNの拡散の場合は48b
BSAの拡散の場合は16hまたは24bインキュべ-卜す
る。
⑦膜からo.1%FBS/M199を抜き取り、
long/ml
TGF一β 1/0.1%FBS/M199
を300〃1加え、プレインキュベ-卜する。またupperとlower液面
が同一になるように、
lowerのゲル上部にM199を100〃1ほど加え
て調節する。
⑧ APPENDIXA-5-3で培養した6穴プレートから培地を吸引し、
PBS
を加え洗浄する。
⑨ 10%trypsin仲BSを1ml加え、細胞がはがれているか顕微鏡で確認す
る。
⑩セルスクレイパーで細胞をはがし、
3穴分をまとめて15ml遠心管
に移す。
⑪ 750rpm/5分遠心分離を行う。
⑫上澄み液を吸い取り、
1mlの0.1%FBS仇4199を加え、細胞懸濁液を
作成する。
⑬血球計算盤で細胞数をカウントし、
l穴当り細胞数が5万個となる
ようにupperに播種後、軽くピペッティングを行う。
㊨ 72h培養する。
A-7-5.遊走細胞数の計測
<使用機器・器具>
・撹拝遠心機(LABINCO/2790001)
・血球計算盤
・オリンパス倒立型顕微鏡(OLYMPUS/CK2)
S-1
Collagenase
(Wako/038-10531)
・
・
・
・
M199
PBS
(SIGMA/M2154)
(SIGMAD8537)
TRYPSIN-EDTA
SOLUTION
(lox)
(SIGMA/T4174)
・ディフ・クイック(シスメックス/16920)
:_有人/?I: )(デニ院
tL.J、;三:研'光村
100
<操作>
I
upper側
①
APPENDIXA-5-4で培養したトランスウェルのupperから培地を吸
い取り、
②
15ml遠心管に移す。
750rpm/5分遠心分離を行う。
上澄み液を吸い取り、 70〃1程度のM199を加え、細胞懸濁液を作
③
成する。
※上澄み液の吸い取ったあとに残った液量によって、細胞懸濁液量が
多少異なってしまうため、この際にピペットなどで全容液量を確認
しておくこと。
④
Ⅲ
血球計算盤にて細胞数を計測する。
膜上、膜下
①膜上、膜下に10%trypsinnBS溶液に浸し、
10分間インキュベ-ト
をする。
②プレートを軽くゆすり、数回膜-ピペッティングを行い、膜上、膜
下の液をそれぞれ15ml遠心管に移す。
③ 750rpm/5分遠心分離を行う。
④上澄み液を吸い取り、
70〃1程度のM199を加え、細胞懸濁液を作
成する。
※上澄み液の吸い取ったあとに残った液量によって、細胞懸濁液量が
多少異なってしまうため、この際にピペットなどで全容液量を確認
しておくこと。
⑤血球計算盤にて細胞数を測定する。
Ⅲ膜内
①ディフクイック固定液に5回/秒浸す。
②ディフクイック染色液Ⅰに5回/秒浸す。
③ディフクイック染色液Ⅲに5回/秒浸す。
④ CAMEDIA
(200倍)で染色された細胞数を数える(10視野)
※染色液に浸す時間及び回数は染色の度合いを見て、調節すること0
ⅠV gel
① lower内collagengelをピペットチップでゆっくりとかき回し、細か
い断片にわける。
② collagenaseS-1を40〃1
(最終濃度がo.o2%となるように、すなわち
101
ゲル1mlに対して100〃1)を加え、溶液が全体に浸透するように再
びピペットチップでかき混ぜる。
③ 37℃で30分インキュベ-卜する。
④ lowerからピペットを用いて液を吸い、
15ml遠心管に移す。
⑤ 750rpm/5分遠心分離を行う。
⑥上澄みを吸い取り、
M1991mlを加えて、軽くvortexをかける。
⑦ 750rpm/5分遠心分離を行う.
※⑥、 ⑦の操作を2、 3回線り返し、細胞を洗浄する。
⑧上澄み液を吸い取り、
M199
70〃1程度加え、細胞懸濁液を作成す
る。
※上澄み液の吸い取ったあとに残った液量によって、細胞懸濁液量が
多少異なってしまうため、この際にピペットなどで全容液量を確認し
ておくこと。
⑨血球計算盤にて細胞数を測定する。
F;i
:.車人'';・r:大ノ、i::
l■.
'、;,I:研究村
102
A-8.細胞増殖試験
<使用機器・器具・試薬>
6穴プレート(住友ベークライト/NS80060)
・
M199培地(SIGMA仇蛇154)
・
(SIGMA/F9423)
FBS
・
1
TGF-β
・
(三光純薬/341635)
FN
・
(SIGMA/T7039-2-UG)
・デジタルカメラCAMEDIAC-3040ZOOM
(OLYMPUS/CK2)
・位相差顕微鏡cK2
<操作>
(丑6穴プレートに10%FBS/M199を2mlずつ加え、プレインキュベ-卜
する。
② 75cm2にcon且uentに培養したHPMCを継代作業し、細胞懸濁液を作
成する。
③血球計算盤にて細胞数をカウントし、5×104個/1穴となるように6穴
プレートに播種する。
④播種後24h培養し、細胞を接着させる。
⑤培地を吸引後、
o.1%FBSバM199を2ml加え24h培養し、
Growth
arrest
を行う。
⑥培地を吸引し、以下の4種類の条件の培地をそれぞれ2ml加える。
・
・
・
・
10%FBS仙I199
0.1%FBS瓜4199
0.1%FBS/M199+
long/ml TGF-β
1
0.1%FBS/M199+
long/mlTGF-β
1
+
10Jlg/mlFN
⑦位相差顕微鏡で確認しながら注射針を用いて、それぞれの条件のプレ
ート底面に3箇所ずつ印(傷)をつける。
24、
48、
72、 96hごとに印を付けた同一部位を、デジタルカ
⑧培養o、
メラにて撮影を行う。
⑨撮影した画像データから細胞数をカウントする。
A-S-1.細胞数のカウント(ImageJ)
① 動画解析同様に、
①を行なう
103
②
③
「Plugins」を選択し、
「cell_co11nter」
cellcounterのツールバーから、
「Ce11counter」を選択する
-
「Initlalize」
-
「Typel
-8」を選択し、細胞をク
リックするo
④クリックしていくと,
「Typel-8」の横に細胞数が表示される.
*細胞がはっきりして見やすいようなら、動画解析と同様に処理を行なうことですばや
く細胞教を測定することができるo
(cellcounterを使う必要がない)
三T,:大I'アニ人乍院
L7:
M- '兜f-Il一
104
B-I.滅菌について
<高圧蒸気滅菌>
高圧蒸気滅菌(オートクレーブ)は、滅菌したい器具を専用の滅菌缶
や金属缶、ケースなどに入れ、滅菌テープで封印し121℃/20分にて行う。
滅菌後は50℃のオーブンに入れ、乾燥・保管する。
液体の高圧蒸気滅菌は専用の耐圧ビンに入れて行う。但し、液体は全
体が均一な温度(121℃)になるまで時間が掛かるため、通常の器具滅菌
の場合よりも1Lあたり20分程度余分に行う。
<乾熱滅菌>
乾熱滅菌も滅菌したい器具を専用の滅菌缶や金属缶に入れて行う。ピ
ペットの場合は吸入口に専用の綿を詰め、滅菌缶に入れて滅菌する。そ
の他のガラス器具などは口にアルミホイルで蓋をし、そのまま滅菌する。
乾熱滅菌の温度と時間は以下の通り(Table.B-1-1)。
Table.B-111乾熱滅菌
一般的な滅菌
160-170℃/2-4h
もしくは
180-200℃/0.5-1時間
RNase丘eeにするための滅菌
180℃/8時間以上(oVerNigbt)
もしくは
250℃/0.5時間以上
※乾熱滅菌は冷めるまでが滅菌である。必ず冷めるまで待つこと。
※高圧蒸気滅菌と乾熱滅菌のいずれの場合も、必ず滅菌対象の材質・
耐熱温度を確認すること。機器の損傷の恐れもある。
<ガス滅菌>
高温耐性のない器具を滅菌する際に使用する。滅菌対象物とガスのバ
イアル瓶を専用の袋に入れ、密封後にガスのバイアル瓶を折り、ガスを
充満させる。このガスは人体にも有害であるため、作業は必ずドラフト
内で行い、開始後2週間程度ドラフト内に放置する。
<uv殺菌>
クリーンベンチは使用後に必ずuv殺菌灯を点灯し、ベンチ内を殺菌
する。その際、ベンチ内のプラスチック製品はアルミホイルで巻くこと。
105
<主要器具の滅菌方法>
○クリーンベンチ内器具一式
×1
・ピンセット
・
t
50ml、
100mlビーカー
シャーレ
×各1
×2
①
ブラッシングにより洗浄後、水洗いし、蒸留水で軽くリンスする。
②
滅菌缶に器具を詰め、滅菌テープで封印し121℃/20分にて高圧蒸
気滅菌を行う。
○手術器具一式
・ハサミ(直、先丸)
・紺子
(拘無し)
×1、
・ピンセット(拘つき)
×3-4
×各1
×1
×20
・滅菌糸(20cm程度のもの)
専用の金属トレイに器具を詰め、滅菌テープで封印し121℃/20分
にて高圧蒸気滅菌を行う。
○チップ、
ReactionTubewithCap
(pcR専用チューブ)
専用のケース、金属缶に詰め、滅菌テープで封印し121℃/20分に
て高圧蒸気滅菌を行う。詰める作業は必ず手袋を着用して行うこと。
0
10mlピペット、パスツールピペット
① 20倍希釈の無りん洗剤に約24時間浸漬後、約24時間水洗いする
(浸漬液は2ケ月毎に交換する)0
② 水気を切り、
③
10mlピペットは吸入口に綿栓を、パスツールピペッ
トは綿栓をせず、 3本毎にアルミホイルで巻く。
滅菌缶にピペットの種類毎に6割程度詰め、180℃で4時間乾熱滅
菌する。
○ゲル作成用三角フラスコ、メスシリンダー
①
ブラッシングにより洗浄後、水洗いし、蒸留水で軽くリンスする。
②
乾燥後、口に7cm四方程度のアルミホイルで蓋をする。
③
そのまま180℃/OverNightにて乾熱滅菌を行う。
:.車人乍人J7:院
卜J';':LTJ)F究村
106
B_2. Primer
List
本研究にて使用したPrimerの詳細を以下に示す。
TableB12_1
List
Primer
Size
Primer
Sequence
Cycle
(bp)
Sense
5'-GATGTCATCATAmGGCAGGTT-3'
GAPDH【49]
antlSenSe
Sense
antisense
5'-GCTTGTGGAGCAGGGACATTC-3'
Sense
5'-TCCCÅrCAGCTGCCCAGAAA-3'
E-ca曲erin【50】
Sense
a5-integrin[51]
antisense
Sense
38
4$7
36
500
39
453
29
292
5'-TGACTCCTGTGTTCCTGTTA-3'
5'-GGCATCAGAGGTGGCTGGAGGCTT」3'
5'-GGCAGCTÅTGGCGTCCCACTGTGG-3'
5'-ACAGCCTGGCÅrGGGGCAAGT-3'
MMP-2【57]
antisens¢
322
5'-TCAGACGAGGACAGTGGGAAAG-3'
snai1【50】
antlSenSe
29
5'-CCTGCACCACCAACTGCTTAGCCC-3'
5'-TTCTCCTCCÅTCCAGTGGAG-3'
※ primerの作成はInvitrogen株式会社に外注した。
107
B-3. RT
/ PCR
program
本研究にて使用したRT及びpCRprogramの詳細を以下に示す。
< RT
program
>
RTprogram
Table.B-2-3
Program
RTprogram
01
Step
Temp/Time
1
95.0℃/5min
Hold
4.0℃/20.0℃/20
1
mln
RTprogram
42.0℃/40
2
02
< PCR
program
min
3
95.0℃/5min
Hold
4.0℃/-
>
Table.B-2-4
PCRprogram
Cyc1
Primer
Step
Pre-PCR
PCR-cycles
denature
94.0℃/3min
94.0℃/30sec
annealing
60.0℃/1min
60.0℃/1min
Post-PCR
eS
GAPDH
29
extension
94.0℃/40sec
60.0℃/10
mln
72.0℃/1.5
72.0℃/1min
min
Hold
denature
Snail
38
4.0℃/95.0℃/3min
94.0℃/1min
annealing
58.0℃/1min
extension
72.0℃/lmin
72.0℃/10
min
Hold
E-cadherin
36
denature
annealing
A__▲______ー_
4.0℃/95.0℃/3min
94.0℃/1min
58.0℃/1min
コ与^Or1/....;..
卜J';I:桝究i・:E・
-A)Tl:人ノデニ人・L-if二P;.二‡
′T′ヽ′ヽOrl/1′ヽ
108
EEZZl
Hold
4.0℃/-
denature
(5-integrin
95.0℃/SOB
95.0℃/30s
annealing
58.0℃/So§
extension
72.0℃/30s
36
72.0℃/7min
Hold
denature
MMp-2
4.0℃/94.OoC/5min
94.0℃/1min
extension
60.0℃/1min
29
annealing
72.0℃/lmin
bold
4.0℃/-
※ 出展はPrimerと同様
DEPC処理水の作成
① Milli-q水を密閉できるガラスビン(青蓋の500mlメディウムビン)
に入れる。
② 500mlのMi11i-q水に対しDEPCを500〟l
(0.1%)加え、蓋をきっ
ちり締めてよく振り混ぜる。
※ DEPCは発癌性があるため、この作業は全てドラフト内で行うこと。
③そのまま37℃のオーブン内に入れ、
2時間加温する。加温の途中に
何回か振り混ぜること。
※振り混ぜることでビンの蓋や首の部分、液体内にDEPCが均一に広
がり、全体のRNaseを失活させる。
④ ビンの蓋を十分緩めて、
121℃/40分(1本あたり500mlの場合)高
圧蒸気滅菌にかける。
※高圧蒸気滅菌をすることによりDEPCを除去する。滅菌後にもDEPC
の臭いが残っている場合は、もう一度高圧蒸気滅菌を行う。
※ DEPCは発癌性であるため、むやみに蒸気を吸わないよう気を付け
ること。また、滅菌装置内に溜まった水は、気化して抜けたDEPC
が溶け込んでいるので、その排水は責任を持って処理すること(大
量の水と共に排水口-)。
滅菌水の作成
①
脱イオン水を密閉できるガラスビン(赤蓋のILメディウムビン)
109
に入れる。
②
蓋をきっちり締めた後、滅菌テープで封印し121℃/40分にて高圧蒸
気滅菌を行う。
B-4.統計処理
<使用機器・器具・試薬>
I
Slat Ⅵew
fわr Windows(vcr.5.0/SAS
Institute
lnc.)
<操作>
① statⅥewを起動したo
② 「ファイル」の「新規」を選択し、以下のように入力したo
チり1
さ-1J「:
エリこ
†芋ち
軍SI
ユ一ワ-)..71
ユー-TT-フ..71
名藁葺&乍N
1重信‡古今[:
軍a
1重吉_
[司冨.J.;ヤ占自由持主it: 風.
・}
ー 〔O㌻.I.r.:.1
1.1;[l:]Ll
1l;l[llー
(I:■ntr :)
1[LLIEJIIu⊂亡.Ee
A
.._q8き
EItW
1l川T.L(.1主lll√(.-:
l
u
i
1
サンプル名を入力
Fig.B-1
Stat
測定値を入力
Ⅵew記入例
③ 「解析」の「新規ビューシート」を選択し、変数一覧の列1、列2をダブ
ルクリックした。
㊨ 2群の比較(対応なし)を選択し解析実行した。
⑤ oK.をクリックし、
P値を求めたo
J、
〉
l
A;・'tlr・≡
110